Production of Hybrids between T. aestivum L. and Ae. ventricosa Tausch
Production of Hybrids between T. aestivum L. and Ae. ventricosa Tausch

Abstract
The purpose of the current study was to estimate the crossing ability and chromosome pairing behavior of intergeneric F1 hybrids between Ae. ventricosa (DvDvNvNv) and Triticum aestivum (AABBDD) crosses. During the production of T. aestivum – Ae ventricosa hybrids, it was observed that F1 hybrids' morphological traits were intermediate between parents, the plants grew vigorously but were completely sterile. Seed set ranging from 2.17 to 8.00 %. Hybrids (2n = 5x = 35) produced without embryo rescue methods. The present study revealed that, the average frequency of chromosome associations at metaphase I was averaged 25.39 univalents and 4.78 bivalents. All received F1 hybrid plants were identical, they exhibited good tillering ability and manifested traits from both parents.
1. Introduction
The tetraploid species Aegilops ventricosa Tausch (syn. Triticum ventricosum) belongs to the tribe Triticeae. It is closely related to wheat (Triticum aestivum L.) (2n = 6x = 42, AABBDD) since its genomic composition is DvDvNvNv, with the Dv and Nv genomes partially homologous to the D genome of Aegilops tauschii L. and to the N genome of Aegilops uniaristata Vis., respectively
. It carries many valuable genes, such as resistance to eyespot , cereal cyst nematode , , Hessian Xy , leaf, yellow and stem rust , , and karnal bunt . Mapping of the rust resistance genes Yr17, Lr37, and Sr38 using molecular markers revealed the presence of a small translocation between the Ae. ventricosa chromosome 2Nv and chromosome 2AS of bread wheat .Dosba et al. (1978) produced a set of wheat – Ae. ventricosa disomic addition lines
. The added pair in one of them was easily identified in mitotic preparations because it was subtelocentric . Both its sporophytic and gametophytic compensations for 6D were high, so the initial expectation was that the Ae. ventricosa pair would prove to be fully homoeologous with wheat homoeologous group 6. It was thus designated as 6Nv.Genetic recombination occurs when the parental genomes share enough homology for homoeologous chromosomes to pair and recombine during meiosis. Chromosome pairing in the interspecific hybrid may occur in two forms: (1) autosyndesis, pairing occurs between members of the same parental genome, and (2) allosyndesis, pairing occurs between members of different parental genomes
. If chromosomes behave as univalents, do not pair during meiosis, or pair autosyndetically DNA is not exchanged between genomes and introgression through genetic recombination will not be possible. A requisite amount of homology between the different genomes will allow homoeologous chromosomes to recombine during meiosis, resulting in recombinant chromosomes carrying both donor and recurrent parent genetic material. All forms of introgression have been successful in transferring agronomically useful traits from wild relatives into cultivated species . Interspecific hybridization can greatly increase the amount of genetic variation available to plant breeders for improvement.The aim of this study was to investigate the crossing ability and morphology of wheat – Ae. ventricosa hybrids and their meiosis behaviour.
2. Material and methods
For the material of the study 2 common wheat lines were used – 171 and 172ACS ({Aegilotriticale [(T. durum Desf. × Ae. tauschii Coss.) × Secale cereale L. ssp. segetale Zhuk.] × T. aestivum L. ‘Chinese Spring’, N500 [(T. aestivum var. graecum Suriya × Ch. Spring) × cv. Bezostaya-1], and 2 Ae. ventricosa accessions.
The emasculation and pollination in the field were carried out during the months of April-May in 2019 at the Absheron Research Base of Genetic Resources Institute. No embryo rescue or hormone treatment was applied for the production of F0 seeds. The spikes were harvested and the number of seed-set per spike was counted about 20 days after pollination. Crossability was expressed as the percentage of the number of seed set to the total number of florets pollinated.
For meiotic studies, young spikes at the appropriate stages were fixed in Carnoy’s fixative for 24 h and then stored in 70% ethanol. Pollen mother-cells were stained and squashed in 1% acetocarmine. Meiotic observations were made and documented with Motic (China) microscope.
3. Results
Table 1 - Crossing ability of Ae. ventricosa with T. aestivum
№ | Cross combinations | Seed setting, % | Obtained seed, absolute number | Plant, absolute number |
1. | 171ACS × Ae. ventricosa | 2.17 | 1 | - |
2. | 171ACS × Ae. ventricosa TA-1999 | 6.94 | 5 | 3 |
3. | 172ACS × Ae. ventricosa TA-1999 | - | - | - |
4. | Ae. ventricosa × 172ACS | 8.00 | 4 | - |
5. | 172ACS × Ae. ventricosa | 4.35 | 2 | 2 |

Spike of hybrid 171ACS × Ae.ventricosa (center), bread wheat parent (left) and wild parent (right)
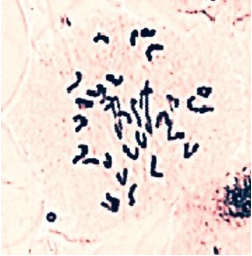
Meiotic behaviour of F1 plant
Reciprocal crosses carried out between 172ACS and Ae. ventricosa. Seed setting of combination when Ae. ventricosa TA-1999 used as a female parent was 8.00%, however obtained 4 seeds did not germinate. Seed setting of 172ACS × Ae. ventricosa combination was 4.35%. Obtained 2 seeds germinated and gave hybrid plants that had intermediate morphotype. The height of the plants was about 103 cm and were completely sterile. Thus, it was not possible to get any seed from 1892 spikelet flowers. During the study of meiosis process of hybrid plants, the number of ring and rod bivalents for each PMCs was 1.75 and 2.89, respectively, the number of univalent was 25.60, the amount of trivalents 0.04 and chiazma frequency was approximatedly 6.47.
Backross hybridizations between F1 plants and different bread wheat lines (171ACS, N500, cv. Siete cerros and cv. Zmitra) was failure.
Kihara (1949) reported the three plants of the cross "Courtdt" × Ae. ventricosa I had 2n = 34 chromosomes (5B absent) and, of those resulting of the crosses "Chinese Spring" × Ae. ventricosa II, one had 34 chromosomes and the other two 35 chromosomes. They grew quite normally and were intermediate to their parents in most morphological characters. The hybrid plants were sterile, and the seed setting was very low
. These results were similar with ours.Table 2 - Study of the meiosis process in F1 hybrids between common wheat lines and Ae. ventricosa
Hybrid combinations | ATH, absolute number | Bivalents, absolute number | Ring bivalents, absolute number | Rod bivalents, absolute number | Univalents, absolute number | Trivalents, absolute number | Chiazma frequency,absolute number |
171ACS × Ae.ventricosa | 142 | 4.91±0.45 | 0.54±0.11 | 4.37±1.46 | 25.18±0.85 | - | 5.44±6.58 |
172ACS × Ae. ventricosa | 97 | 4.64±0.28 | 1.75±0.40 | 2.89±1.78 | 25.60±0.88 | 0.04±0.22 | 6.47±0.91 |
4. Discussion
Obtaining interspecific hybrids offers significant variability. It is the crucial step in any program of genes introgression from wild species. However, its success depends not only on the choice of genitors that suits the objectives of the program, but also on crossing affinity.
Previous works had shown the effectiveness of a method to transfer genes from the wild grass Ae. ventricosa to the cultivated wheat T. aestivum
, . The strategy involves a cross of the donor species, Ae. ventricosa with T. turgidum (AABB), which acts as a bridge, followed by the rescue of the sterile ABDvMv hybrid with pollen from the recipient species T. aestivum (AABBDD). Plants from this cross were fertile and stable lines with 42 chromosomes were derived from them after repeated selfing.The few works dedicated to interspecific crosses where species of the genus Aegilops are taken as female parent, report the weakness of obtaining such hybrids, which seems, more favourable in field conditions . Many criteria influence the acquisition of fertile hybrids and backcross progenitor for introgression between two genera, including genetic relationships, ploidy level, and hybridization direction . The success of obtaining hybrids depends largely on the parental genotypes involved in the crossing. The differences in hybridization affinity of Aegilops species and wheat varieties are highly observed , , . In many studies, the common sharing of the D genome between the bread wheat and the wild parent allowed the pairing of homeologous chromosomes and obtaining fertile hybrids , .
The distribution of biochemical markers and the maximum number of chromosomes from the Mv genome in the selected lines obtained by Olmedo
. The latter is deduced from the number of univalents at MI observed in the hybrid and represent an overestimation for two reasons:I) because it has been repeatedly observed that there is incomplete homology between the D genomes of T. aestivum and Ae. ventricosa, and
II) because the transfer of a terminal segment of a chromosome from the Mv genome to a wheat chromosome would suffice to drastically reduce its meiotic pairing with the original wheat chromosome.
It can be concluded, from the joint consideration of the biochemical and the cytological data, that the genetic transfer from the Mv genome of Ae. ventricosa to hexaploid wheat has taken place not only by chromosome substitution but also by recombination. In other words, lines carrying, at most, one whole Mv genome chromosome also carry two independently inherited Mv genome markers.
E. D. Badayeva informed that hybridization of species that share common genomes can induce an extensive exchange of genetic material between closely related chromosomes. According to meiotic analysis, the D-genome of Ae. ventricosa is only slightly modified relative to its diploid progenitor
. The D-genome of common wheat is also very similar to Ae. tauschii. We might expect that wheat–Ae. ventricosa D-genome chromosome pairing could have occurred in the F1 hybrid plant, giving rise to recombinant D-Dv genome chromosomes. Indeed, a high level of pairing was observed in wheat–Ae. ventricosa hybrids . In contrast, the Nv genome is quite distant from the A- and B-genomes, and they rarely pair at meiosis . These results underline the use of complementary markers, such as biochemical, molecular, and cytological markers in the production and characterization of addition and introgression lines, particularly if the parental species share a common genome which interferes with meiotic pairing in hybrids.5. Conclusion
At interspecific hybridization between bread wheat genotypes and two accession of wild relative, Aegilops ventricosa was achieved with low crossability rate for all genotypes. Here presented results are only the initial step of the involvement of wild species Ae. ventricosa in long process of production of bread wheat breeding lines with introgressed alien genes. Successive progenies are going to be screened at morphological, physiological, cytological and molecular level for hybrid identification and enhancing of genetic variation for biotic – and abiotic stress resistance traits and its incorporation into common wheat. All wheat-aegilops hybrids involved backcross hybridization, but it was ineffective.