DYNAMICS OF LIVE GROUND COVER OF PINE AND SPRUCE CROPS DEPENDING ON GROWTH CONDITIONS
DYNAMICS OF LIVE GROUND COVER OF PINE AND SPRUCE CROPS DEPENDING ON GROWTH CONDITIONS
Abstract
The structure of the herbaceous layer of forest cenoses, its quantitative and qualitative composition are largely determined by the microclimate of the environment, which is formed and changed during the growth and development of forest crops. Observations of forest crops in different growth phases visually illustrate the dynamics of the ongoing succession of plant communities.
The aim of the research is to study the dynamics of species diversity of the live ground cover of forest crops depending on their species composition and age, as well as growth conditions.
Monitoring studies of the live ground cover of forest crops were carried out. As a result of the research it was concluded that after anthropogenic disturbance of phytocenoses there is a transformation of ecological and cenotic structure of flora in the direction of restoration of the original structure of forest environment. The nature of changes occurs along the path of gradual displacement of meadow species by the active forest group of plants, as well as by reducing the proportion of less stable inactive and low-active species.
1. Введение
Изучение растительности нижних ярусов на лесокультурных площадях играет важную роль в исследовании скорости лесовозобновительного процесса. Напочвенному покрову в фитоценозе отведена значимая ценотическая роль: растения способствуют увеличению мощности плодородного горизонта почвы, повышают активность микрофлоры и зоокомпонента почвы и др.
В связи с этим необходимо всестороннее изучение растений напочвенного покрова, определение наиболее активных видов, которые непременно влияют на ход лесовозобновительных процессов.
Применительно к лесохозяйственной практике предлагаем принять за основу активности видов – способность растений участвовать в формировании и смене растительных сообществ с помощью многообразных механизмов адаптации, которые в своей совокупности помогают преодолевать негативное воздействие неблагоприятных факторов, выживать, размножаться и быть устойчивыми в фитоценозах. Классификация активности видов является теоретической основой оценки антропогенной нагрузки в ценозе, а также служит сигналом для выявления значимых видов.
2. Объекты и методы исследований
Тест-объектами наших исследований явились культурфитоценозы антропогенно-природного происхождения сосны (Pinus sylvestris L.) и ели (Picea abies (L.) H. Karst.) в фазе смыкания (14 лет) и чащи (22 года), то есть до формирования исконно лесной среды. Объекты исследований расположены на территории Архангельской области.
В работе использовали общепринятые в лесоводстве и таксации методики исследований древостоя .
Количественный анализ флоры или парциальная активность видов (ПарцА) оценивалась на основании методических подходов Б.А. Юрцева . В основу фитоценотической классификации активности видов растений (ФитоА) по широте эколого-ценотической амплитуды легли геоботанические исследования , , , . На основе проведенного анализа методических подходов к определению активности видов в работе была предложена и использована адаптированная классификация активности растений напочвенного покрова культурфитоценозов в условиях северной части Восточно-Европейской равнины. Усредненная таксационная характеристика исследуемых древостоев представлена в таблице 1.
Таблица 1 - Усредненные таксационные показатели культур
Вариант | Густота, тыс. шт./га | Средний диаметр, см | Средняя высота2, м | Класс бонитета | Полнота | Запас древесины, м3/га | |
первоначальная | на момент исследования | ||||||
Кисличный тип лесорастительных условий Дренированные условия в возрасте1 14 лет | |||||||
Сосна | 2,5 | 2,2 | 8,3 | 6,3 | I | 0,8 | 69,9 |
Ель | 3,5 | 2,9 | – | 1,8 | V | – | – |
22 года | |||||||
Сосна | 2,5 | 2,1 | 12,6 | 11,3 | Iа | 1,1 | 164,7 |
Ель | 3,5 | 1,1 | 6,7 | 5,3 | IV | 0,4 | 18,1 |
Черничный тип лесорастительных условий Условия временно избыточного увлажнения в возрасте1 14 лет | |||||||
Сосна | 2,5 | 1,6 | 6,9 | 5,7 | II | 0,4 | 24,7 |
Ель | 2,8 | 2,4 | – | 1,8 | V | – | – |
22 года | |||||||
Сосна | 2,5 | 1,4 | 11,2 | 9,8 | I | 0,6 | 70,3 |
Ель | 2,8 | 2,1 | 5,1 | 4,5 | IV | 0,4 | 16,1 |
Примечание: 1 - с момента создания культур; 2 – при средней высоте древостоя менее 3 м в качестве ведущего показателя принята высота культур в соответствии с рекомендациями В.В. Огиевского, А.А. Хирова [6]
Ландшафтная активность (ЛандА) определена путем суммированных парциальных оценок с учетом широты эколого-ценотической амплитуды:
где ЛандАi – ландшафтная активность i-го вида (балл), ФитоАi – балл фитоценотической активности i-го вида, ПарцАi – балл парциальной активности i-го вида, Pi – балл постоянства i-го вида, Oi – балл обилия i-го вида, N – число описаний.
3. Основные результаты
При выполнении сравнительного анализа флористических показателей напочвенного покрова, исследованных нами культурфитоценозов, произрастающих в разных условиях местопроизрастания, следует отметить, что для насаждений черничного типа леса в условиях временного избыточного переувлажнения значения показателей видового богатства травянистых растений ниже, чем в кисличном типе леса на дренированных почвах.
Такой характер развития живого напочвенного покрова связан с особенностью временно переувлажненных экосистем, которые имеют более высокую влажность в сравнении с дренированными почвами. Избыточное увлажнение ведет к изменению температурного режима почв, снижению ее аэрации и, как следствие, уменьшению объемной массы и повышению кислотности. Такое сочетание факторов среды ведет к формированию природных условий, в которых может существовать строго ограниченный видовой состав растений.
В кисличном типе леса к моменту смыкания культур сосны и ели в живом напочвенном покрове выявлено 36 и 33 вида растений соответственно, в том числе 2 вида кустарничков (Vaccinium vitis-idaea L., Vaccinium myrtillus L.), 2 вида моховой растительности (Polytrichumcommune Hedw., Pleuroziumschreberi (Willd. Ex Brid.) Mitt.) и 2 малолетних травянистых вида (Melampyrum pratense L., Campanula patula L.) (рис. 1).
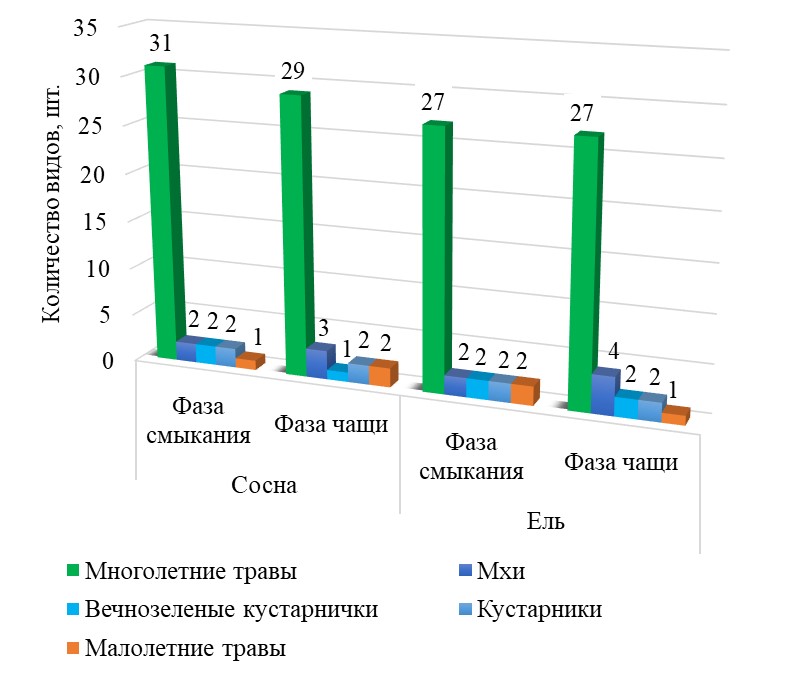
Рисунок 1 - Видовое богатство растительных сообществ культур сосны и ели на дренированных почвах
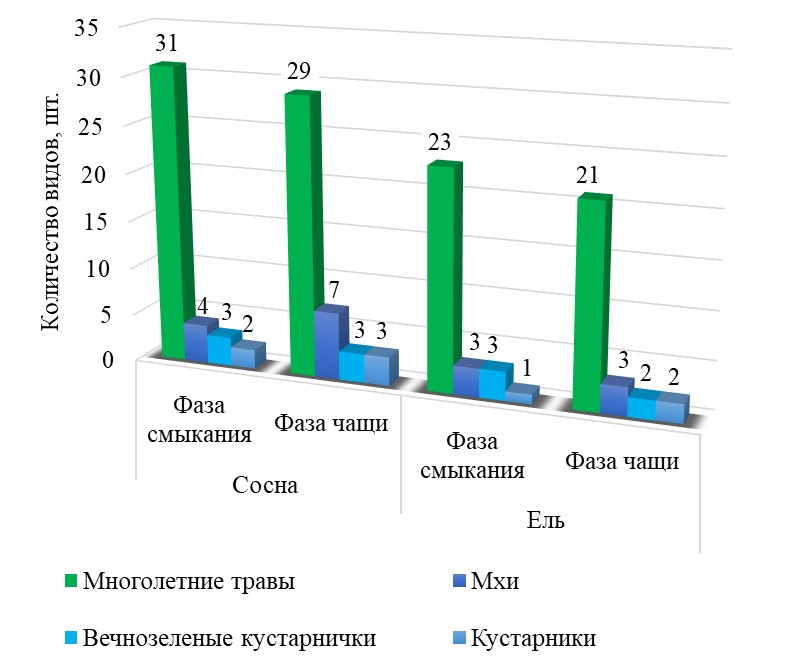
Рисунок 2 - Видовое богатство растительных сообществ культур сосны и ели в условиях периодического переувлажнения почв
Относительное участие видов разнотравья и кустарничков на всех исследуемых участках с момента смыкания древостоя до достижения ими фазы чащи постепенно снижается в диапазоне от 12 до 45%.
Такой зрительный эффект упрощенности структуры сообщества, снижение видового разнообразия при движении к климаксу свойственны таежной зоне . По мнению ряда авторов, внутренняя структура ценопопуляции усложняется только за счет возрастной дифференциации, а структура сообщества в целом – за счет мозаичности , , , .
Проведенный нами анализ доказывает возрастные изменения видового состава растительных сообществ. К фазе чащи происходит постепенное уменьшение общего числа видов, при этом существует достоверная (p = 0,05) высокая связь между общим числом видов и возрастом лесных культур (r = -0,77).
Как показывают наши данные, флористический состав живого напочвенного покрова с возрастом культур существенно видоизменяется. На смену луговым растениям, которые характерны для открытых пространств, приходят типично лесные виды. Изменения в составе луговой растительности особенно заметны на примере дренированных почв, поскольку на данном этапе формирования культур древесный ярус здесь отличается более высокими таксационными параметрами (табл. 1).
К 22-летнему возрасту лесных культур с момента создания мхи увеличились в видовом количественном отношении в 1,8 раза: отмечены новые виды Hylocomium splendens Hedw., Plagiomnium undulatum (Hedw.) T.J. Kop., Ptilium crista-castrensis (Hedw.) De Not., Rhytidiadelphus triquetrus (Hedw.) Warnst.
Как нам кажется, оценка флористического состава недостаточно полно характеризует ход сукцессии растительных сообществ. В этой связи для характеристики богатства и изменчивости живого напочвенного покрова лесных культур мы использовали классификацию активности видов. Она качественно дополняет флористический анализ, тем самым позволяет оценить иерархические позиции видов в растительном сообществе (табл. 2.).
Таблица 2 - Систематическая структура растительных сообществ на участках культур сосны и ели
Семейство | Количество видов в условиях | |||||||
дренированных почв | временного переувлажнения | |||||||
сосна | ель | сосна | ель | |||||
фаза смыкания | фаза чащи | фаза смыкания | фаза чащи | фаза смыкания | фаза чащи | фаза смыкания | фаза чащи | |
Неактивные виды | ||||||||
Бобовые Fabaceae | - | 1 | - | - | 2 | - | - | - |
Вересковые Ericaceaе | 1 | - | 1 | 1 | - | - | - | - |
Губоцветные Lamiaceae | - | - | - | - | - | 1 | - | - |
Злаковые Poaceae | - | - | - | - | - | 1 | 1 | - |
Зонтичные Apiaceae | 2 | - | 1 | - | - | 1 | - | - |
Колокольчиковые Campanulaceae | 1 | 1 | 1 | - | - | - | - | - |
Лилейные Liliaceae | 1 | 1 | - | 1 | - | - | - | - |
Лютиковые Ranunculaceae | - | 1 | 1 | - | - | - | - | - |
Норичниковые Scrophulariaceae | 2 | - | 2 | 1 | - | 1 | - | - |
Первоцветные Primulaceae | - | - | - | - | 1 | - | - | - |
Розовые Rosaceae | 1 | - | 1 | 1 | - | - | - | - |
Сложноцветные Asteracea | 2 | - | 1 | - | 1 | - | 1 | - |
Хвощевые Equisetaceae | 1 | - | 1 | - | 1 | 1 | - | - |
Всего видов | 11 | 4 | 9 | 4 | 5 | 5 | 2 | 0 |
Малоактивные виды | ||||||||
Аспидиевые Aspidiaceae | - | - | - | - | - | 1 | - | - |
Бобовые Fabaceae | - | - | - | - | - | 1 | - | 1 |
Гвоздичные Caryophyllaceae | 1 | 1 | 1 | 1 | - | - | - | - |
Гераниевые Geraniaceae | 1 | - | 1 | - | - | - | - | - |
Жимолостные Capryfoliaceae | - | - | - | - | 1 | 1 | 1 | - |
Злаковые Poaceae | 1 | - | 1 | - | 1 | - | - | - |
Зонтичные Apiaceae | - | - | - | - | 1 | - | - | - |
Кисличные Oxalidaceae | 1 | - | - | - | 1 | - | 1 | - |
Кочедыжниковые Athyriaceae | - | - | - | - | 1 | - | 1 | - |
Крапивные Urticaceae | 1 | - | 1 | - | - | - | - | - |
Лилейные Liliaceae | - | - | - | - | 1 | - | - | 1 |
Мареновые Rubiaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Норичниковые Scrophulariaceae | - | 1 | - | 1 | - | - | - | - |
Осоковые Cyperaceae | - | - | - | - | 1 | - | 1 | - |
Первоцветные Primulaceae | - | 1 | - | 1 | - | - | - | - |
Розовые Rosaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Сложноцветные Asteracea | 1 | 1 | - | - | 2 | 2 | 1 | 1 |
Фиалковые Violaceae | - | 1 | - | 1 | - | - | - | - |
Хвощевые Equisetaceae | - | - | - | 1 | - | 1 | - | 1 |
Всего видов | 8 | 7 | 6 | 7 | 11 | 6 | 7 | 4 |
Среднеактивные виды | ||||||||
Аспидиевые Aspidiaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Бобовые Fabaceae | - | 1 | - | 1 | 1 | 1 | 1 | 1 |
Вересковые Ericaceaе | - | 1 | - | 1 | 1 | 1 | 1 | 1 |
Ворсянковые Dipsacaceae | 1 | - | 1 | - | - | - | - | - |
Гераниевые Geraniaceae | - | - | - | - | - | 1 | - | 1 |
Грушанковые Pyrolaceae | - | 1 | - | 1 | 1 | 1 | 1 | 1 |
Зверобойные Guttiferae | - | 1 | - | 1 | - | - | - | - |
Злаковые Poaceae | - | - | - | - | - | 1 | - | 1 |
Зонтичные Apiaceae | - | 1 | - | 1 | 1 | - | 1 | - |
Кочедыжниковые Athyriaceae | 1 | 1 | - | 1 | 1 | 1 | - | - |
Крапивные Urticaceae | - | 1 | - | 1 | - | - | - | - |
Крыжовниковые Grossulariaceae | - | - | - | - | - | 1 | - | - |
Лилейные Liliaceae | - | - | - | - | 1 | - | 1 | - |
Норичниковые Scrophulariaceae | - | 1 | - | 1 | - | - | - | - |
Первоцветные Primulaceae | - | - | - | - | - | 1 | - | 1 |
Розовые Rosaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Сложноцветные Asteracea | - | - | - | - | 1 | 2 | 1 | 1 |
Хвощевые Equisetaceae | - | 2 | - | 1 | - | - | - | - |
Всего видов | 4 | 12 | 3 | 11 | 9 | 10 | 8 | 7 |
Активные виды | ||||||||
Бобовые Fabaceae | 1 | 1 | 1 | 1 | - | - | - | - |
Вересковые Ericaceaе | 1 | - | 1 | - | 1 | 1 | 1 | 1 |
Ворсянковые Dipsacaceae | - | - | - | - | 1 | - | 1 | - |
Гераниевые Geraniaceae | - | 1 | - | 1 | - | - | - | - |
Зверобойные Guttiferae | 1 | - | 1 | - | - | - | - | - |
Злаковые Poaceae | 2 | 1 | 4 | - | - | - | - | - |
Зонтичные Apiaceae | - | 1 | - | 1 | - | - | - | - |
Кипрейные Onagraceae | 1 | 1 | 1 | 1 | - | 1 | - | 1 |
Кисличные Oxalidaceae | - | 1 | - | 1 | - | 1 | - | 1 |
Кочедыжниковые Athyriaceae | - | - | - | - | - | 1 | - | 1 |
Лилейные Liliaceae | - | - | - | - | - | 1 | - | 1 |
Осоковые Cyperaceae | - | - | - | - | - | 1 | - | 1 |
Розовые Rosaceae | - | 1 | - | 1 | 2 | 2 | 1 | 2 |
Сложноцветные Asteracea | - | - | - | - | 1 | - | 1 | - |
Фиалковые Violaceae | 1 | - | 1 | - | - | - | - | - |
Хвощевые Equisetaceae | 1 | - | 1 | - | 2 | 2 | 2 | 2 |
Всего видов | 8 | 7 | 10 | 6 | 7 | 10 | 6 | 10 |
Высокоактивные виды | ||||||||
Бобовые Fabaceae | 1 | - | 1 | - | - | - | - | - |
Гераниевые Geraniaceae | - | - | - | - | 1 | - | 1 | - |
Злаковые Poaceae | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 |
Кипрейные Onagraceae | - | - | - | - | 1 | - | 1 | - |
Розовые Rosaceae | 2 | 2 | 2 | 2 | 1 | 3 | 1 | 3 |
Всего видов | 5 | 4 | 5 | 4 | 4 | 4 | 4 | 4 |
Итого | 36 | 34 | 33 | 32 | 36 | 35 | 27 | 25 |
Согласно фитоценотическому анализу установлено, что основу сообщества лесных культур в фазе смыкания на дренированном участке составляют высокоактивные виды (1 класс активности) (14…15% от общего количества сосудистых видов ценофлоры) – Deschampsia cespitosa L., Calamagrostis epigeios L., Trifolium pratense L., Rubus saxatilis L., Potentilla erecta (L.) Raeusch. Данные виды широко представлены на всей лесокультурной площади и имеют высокий показатель обилия.
С возрастом древостоя в фазе чащи лесных культур, с изменением освещенности и других микроклиматических факторов местообитания, высокоактивные светолюбивые виды напочвенного покрова Trifolium pratense L. и Potentilla erecta (L.) Raeusch. переходят соответственно во 2 и 3 класс активности, а семигелиофитный вид Rubus idaeus L., наоборот, становится высокоактивным видом. Тем самым, к фазе чащи лесных культур общее количество высокоактивных видов сократилось с 5 до 4, при этом на смену Trifolium pratense L. и Potentilla erecta (L.) Raeusch. пришел Rubus idaeus L.
В условиях временного избыточного увлажнения сохраняет свое доминирующее положение Calamagrostis epigeios L., Rubus saxatilis L., а также с учетом обозначенных почвенных особенностей высокую позицию в фазе смыкания занимает Geranium sylvaticum L. и Chamerion angustifolium (L.) Holub., таким образом, эти виды соответствуют 1 классу активности (11…15% от общего количества сосудистых видов ценофлоры).
В фазе чащи древостоя высокоактивные лугово-лесные виды Geranium sylvaticum L. и Chamerion angustifolium (L.) Holub. переходят соответственно во 2 и 3 класс активности, а лесные виды Filipendula ulmaria (L.) Maxim, Rosa acicularis Lindl., наоборот, увеличивают ландшафтное распространение и становятся высокоактивными.
Группу активных видов (2 класс активности) наряду с высокоактивными, мы также относим к наиболее приспособленным к условиям фитоценозов несмотря на то, что они представлены не на всех учетных площадках.
По нашим данным, их доля в популяции составляет 19…40%. Большинство высокоактивных и активных видов относятся к лесной и луговой ценотической группе. Если рассматривать обе группы, то растения 1 и 2 класса активности достаточно многочисленны. На их долю приходится 31…56% популяции, что составляет 10…15 видов растений. Можно отметить, что большинство из рассматриваемых видов сохранились на лесокультурной площади с вейниковой вырубки 1999 г.
Среднеактивные виды (3 класс активности) мы относим к промежуточной группе. Данные виды отличаются более узким диапазоном толерантности к экологическим условиям, так, например, Urtica dioica L. и Dryopteris filix-mas (L.) Scohott. являются более требовательными к почвенному питанию. Наши исследования показали, что по прошествии 14 лет на лесокультурной площади сохраняется от 9 до 30% среднеактивных видов сосудистых растений, а к 22-летнему возрасту их доля возрастает до 29…35%.
Обратный характер сукцессии мы наблюдаем в группе малоактивных и неактивных видов (4…5 классы активности), представители которых встречаются в небольшом количестве на отдельных участках, либо единичными растениями. Малоактивные и неактивные виды не имеют серьезного лесохозяйственного значения для лесных культур, но для экосистемы в целом увеличение пула видов повышает продуктивность системы, улучшает почвенные показатели. В фазе смыкания на данные группы растений приходится значительная часть спектра – 33…53% от общего количества видов ценофлоры. В фазе чащи доля малоактивных и неактивных видов значительно меньше (16…34%) за счет снижения общего видового биоразнообразия из-за высокого распространения активной части флоры.
Таким образом, чем выше участие активных видов в растительном сообществе, тем меньше ресурсов остается малоактивным видам, тем ниже их адаптационный потенциал и общее число видов. Рост участия активных и высокоактивных видов ведет к случайному вытеснению сопутствующих растений. Кроме того, активные виды изменяют условия произрастания для других видов и могут ограничивать их присутствие, исключая из состава ценоза сначала менее устойчивые виды, а затем все более адаптированные до полного их исчезновения.
К малоактивной группе растений напочвенного покрова лесных культур в фазе смыкания отнесены такие временные луговые виды, как Phleum pratense L., Stellaria graminea L., сорно-рудеральные Cirsium arvense (L.) Scop., Urtica dioica L., а также лесные виды Rosa acicularis Lindl., Geranium sylvaticum L., Galium boreale L., Oxalis acetosella L., пока неактивные, но которые спустя десятилетие (в фазе чащи) в большинстве своем займут 2 класс активности.
Со временем в ценофлоре лесных культур фазы чащи исчезло большинство неактивных видов (Achillea millefolium L., Leucanthemum vulgare Lam., Taraxacum officinale Wigg., Gagea lutea (L.) Ker Gawl., Aegopodium podagraria L., Veronica officinalis L.), а им на смену пришли новые виды (Maianthemum bifolium (L.) F.W. Schmidt, Paris quadrifolia L. и др.), не встреченные ранее.
В культурах ели в фазе чащи по причине недостатка освещенности наблюдается существенное сокращение малоактивных и неактивных видов растений живого напочвенного покрова.
4. Заключение
Учитывая вышесказанное, следует подчеркнуть, что происходит постепенное вытеснение луговых видов активной лесной группой растений. В большей степени изменения происходят за счет сокращения менее устойчивых неактивных и малоактивных видов, таким образом, происходит трансформации эколого-ценотической структуры флоры после антропогенного нарушения в направлении восстановления исходной структуры лесной среды.