INFLUENCE OF ENVIRONMENTAL FACTORS AND DGAT1 GENE POLYMORPHISM ON VARIABILITY OF MILK PRODUCTIVITY TRAITS AND FATTY ACID PROFILE OF MILK OF HOLSTEIN BLACK-BREED COWS
INFLUENCE OF ENVIRONMENTAL FACTORS AND DGAT1 GENE POLYMORPHISM ON VARIABILITY OF MILK PRODUCTIVITY TRAITS AND FATTY ACID PROFILE OF MILK OF HOLSTEIN BLACK-BREED COWS

Abstract
The article presents the results of evaluation of the influence of environmental and genetic factors, including allelic polymorphism of DGAT1 gene (diacylglycerol-O-acyltransferase) on performance and milk fatty acids profile in Holsteinised black-breed cows. A total of 310 cows from the experimental farm were analysed. Information on milk composition and productivity was provided by records of monthly control milkings conducted over two years (total number of records n>10000). No deviations in genotype frequency distribution from the Hardy-Weinberg law were detected. SNPs rs109234250 and rs109326954 localized in the DGAT1 gene had predominant K allele (0.6). The fatty acid composition of milk fat was dominated by saturated medium-chain fatty acids. Heritability and genetic correlations of the studied traits correspond to the ways of fatty acids' formation in the animal organism. The highest estimated value of the fixed effect of duration of lactation (DIM) was found for the traits "fat content" and "saturated medium-chain fatty acids content". Estimation of control milking season factor showed maximum positive influence in winter period with increasing estimation of each subsequent lactation. For long-chain and monounsaturated fatty acids, maximum estimates of the effect of season were shown for the spring period and growth up to the third lactation. Polyunsaturated and trans isomers of fatty acids showed distinctive results with the lowest regression coefficients and maximum effect estimates in the first lactation with a decrease in each subsequent lactation. The maximum effect on fatty acid composition of milk fat was identified for allele A of DGAT1 gene with predominance of AA genotype.
1. Введение
По причине особого влияния на потребительские качества получаемой жиромаслянной продукции профилю жирных кислот молочного жира придается пристальное внимание. За последние десятилетия мнения о вреде или пользе молочного жира менялись на диаметрально противоположные. Это связано с тем, что молочный жир представлен в основном насыщенными жирными кислотами (до 65%), что зачастую ассоциируют с риском возникновения сердечно-сосудистых заболеваний, но в то же время, ряд исследователей сообщает о полезных биологических свойствах олеиновой и стеариновой жирных кислот, что способствуют регуляции уровня липидов в крови. И в дополнение молочный жир содержит такие жирные кислоты как линолевая, альфа-линолевая и коньюгированная линолевая (CLA), полезные с точки зрения питания , , , . Кроме непосредственного влияния молочного жира на здоровье человека при его употреблении, интерес к профилю жирных кислот вызывает его влияние на технологические свойства продуктов переработки молочного жира. Так, увеличение ненасыщенных длинноцепочечных жирных кислот, в противовес короткоцепочечным, увеличивает мягкость и текучесть сливочного масла при низких температурах, но может отрицательно сказаться на вкусовых качествах, по причине усиленного окисления продукта, что также влияет на максимальные сроки хранения .
Состав и содержание жирных кислот молока зависит от путей их синтеза. Условно можно выделить несколько путей образования: непосредственно из рациона, синтез в молочной железе, биогидрирование (бактериальная деградация в рубце) и высвобождение из жировых запасов , . При этом каждому типу жирной кислоты соответствует свой путь синтеза, так насыщенные коротко- и среднецепочечные синтезируются преимущественно de novo в молочной железе, тогда как ненасыщенные и длинноцепочечные жирные кислоты поступают из корма . Регуляция указанных процессов может быть связана со множеством факторов влияющих на них, таких как кормление, порода, номер и стадия лактации. Кормление животного отражается на профиле жирных кислот посредством влияния на регуляцию экспрессии липогенных генов и снижения синтеза de novo и увеличению содержания длинноцепочечных жирных кислот, при повышении содержания полиненасыщенных жирных кислот в кормах. Данные колебания в профиле молочного жира проявляются при смене сезона, т.к. являются отражением смены рациона животных, когда зимой преобладают концентрированные корма и силос, а летом свежая трава, богатая ненасыщенными жирными кислотами . С учетом того, что основная доля жирных кислот синтезируется самим животным, то также стоит брать в расчет и такой фактор как генотип животного, а в частности, полиморфизмы генов регулирующие липидный обмен. Фермент диацилглицерин-O-ацилтрансфераза, кодируемый геном DGAT1 является одним из основных маркеров жирномолочности, т.к. катализирует заключительную стадию биосинтеза триглицеридов. Ассоциации данного гена с содержанием молочного жира и жирных кислот показаны рядом авторов в GWAS – исследованиях, как у молочных пород, так и у мясных , , . Таким образом, целью нашего исследования являлась оценка влияния средовых и генетических факторов на изменчивость продуктивности, а также содержание и состав профиля жирных кислот молочного жира у голштинизированных черно-пестрых коров.
2. Материалы и методы исследования
Образцы ушных выщипов (n=310) для генотипирования и пробы молока были получены от животных голштинизированной черно-пестрой породы (опытное хозяйство ФГБНУ ФИЦ ВИЖ им.Л.К.Эрнста, Краснодарский край). Геномная ДНК выделялась с использованием набора «ДНК-Экстран-2» производства НПО «СИНТОЛ» из выщипов ушных раковин согласно инструкции производителя.
Реакцию амплификации проводили методикой ПЦР в «реальном времени», разработанной Ковальчук С.Н. 2017 г. на приборе Bio-Rad CFX96 в оптимизированных условиях .
В исследовании учитывались следующие показатели: суточный удой (кг), массовая доля жира/белка в молоке (%), содержание казеинов (%), жирнокислотный состав молока (г/100г): С14, С16, С18, С18.1, LCFA, MCFA, MUFA, PUFA, SFA, SCFA, TFA. Данные о фракционном составе молока получали посредством индивидуального отбора проб молока во время проведения ежемесячных контрольных доений – 3 раза в сутки (утро-день-вечер) и последующего анализа в лаборатории селекционного контроля качества молока ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста на автоматическом анализаторе CombiFoss 7 DC («Foss», Дания) в период с июня 2020 по июль 2022 гг. В исследовательскую базу вошли 10243 наблюдения за период проведения исследования.
Описательные статистические параметры (среднее арифметическое, ошибка, стандартное отклонение) вычислялись при помощи пакета «Анализ данных» в среде MS Excel 2013. Степень изменчивости признаков оценивали по коэффициенту вариации (Cv). Для расчета наследуемости (h2) использовали отношение генетической вариансы к сумме генетической и остаточной варианс. Выявленные различия считали статистически значимыми при p≤0,05.
Оценка влияния генетических маркеров и средовых факторов на продуктивные качества и жирнокислотный состав молока производилась при помощи математической модели смешанного типа, составленной в рамках методологии BLUP Animal Model. Расчёты осуществлялись в программах семейства BLUPF90 . Модель оценки имела следующий вид:
где yijklm – оцениваемый показатель продуктивности или жирнокислотного состава молока; µ – популяционная константа; Li – фиксированный эффект i-й лактации; β – коэффициент линейной регрессии; DIMj – количество дойных дней; Seasonk – сезон лактации, в который был произведен отбор образца молока (Зима-весна-лето-осень); Gl – фиксированный эффект каждого генотипа; animalm – рандомизированный эффект m-ой коровы (n=10243 наблюдения) имеющий нормальное распределение со средней, равной 0, и вариансой, равной σa2 (Vara ~ 0, σa2); eijklm – эффект остатков модели.
3. Основные результаты
Для каждого животного были идентифицированы все генотипы исследуемого гена DGAT1 (таблица 1).
Частоты аллелей и генотипов генов DGAT1 у голштинизированного черно-пестрого скота опытного хозяйства
Ген | Частоты генотипов | Частоты аллелей | χ2 | p-value | |||
DGAT1 | AA | AK | KK | A | K | 0,03 | 0,98 |
0,16 | 0,47 | 0,36 | 0,4 | 0,6 | |||
В рамках исследуемой выборки голштинизированных черно-пестрых коров определены частоты встречаемости аллелей А (0,4) и К (0.6) гена DGAT1 SNP rs109234250, rs109326954 с частотой гетерозигот (Ho) 0,47. Отклонений фактического распределения частот генотипов от теоретического согласно закону Харди-Вайнберга не наблюдалось.
Преобладание аллеля К гена DGAT1 SNP rs109234250, rs109326954 в популяциях крупного рогатого скота больше характерно для местных и аборигенных пород скота. Так, у аборигенного африканского крупного рогатого скота частота встречаемости аллеля К составляет от 0,77 до 0,929, в то время как у их помесей с голштинской породой от 0,386 до 0,658 , . Аналогичные данные показаны для черного японского мясного скота, где минорным являлся аллель А с низкой частотой встречаемости генотипа АА – 0,056 . Среди отечественных пород скота повышение частоты встречаемости аллеля К принято ассоциировать с прилитием голштинской крови, особенно у черно-пестрой голштинизированной породы , а также с высокой частотой гетерозигот в различных популяциях холмогорской породы от 0,35 до 0,49 , . Для европейской популяции голштинской породы характерно незначительное преобладание аллеля А, с долей гетерозигот – 0,47 , , , , приближенная ситуация показана и для китайской популяции голштинского скота, с тем исключением, что доля гомозиготного генотипа АА и доля гетерозигот были одинаковы – 0,479 и минимальным присутствием генотипа КК – 0,042 .
Из литературных данных также показано преобладание аллеля А гена DGAT1 в иностранных популяциях мясных и молочных породах крупного рогатого скота, при этом сопоставимые данные получены для таких европейских пород скота как симментальская, бурая, шароле, лимузин, абердин-ангуская, герефорд, у которых частота встречаемости аллеля А превышала 0,8, а иногда наблюдалось полное отсутствие аллеля K в популяции , , , .
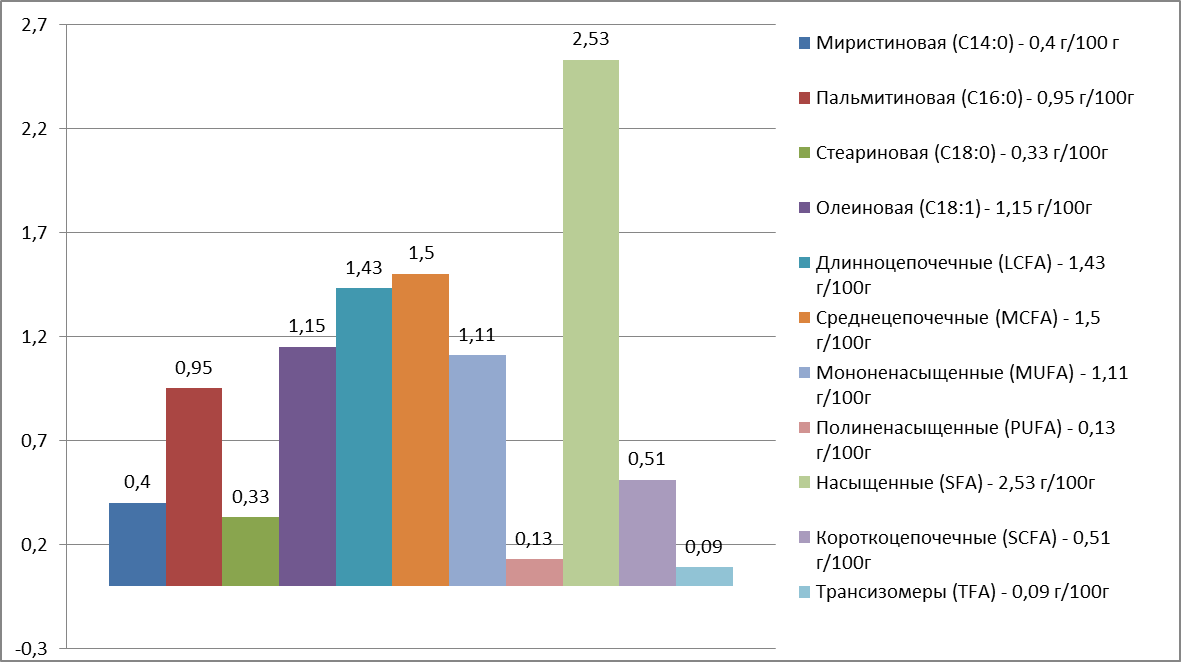
Содержание жирных кислот в молоке голштинизированных черно-пестрых коров опытного хозяйства
Наследуемость показателей продуктивности и жирнокислотного состава молока колебалась в диапазоне от 0,13 до 0,58 (табл. 2). Для длинноцепочечных (LCFA), мононенасыщенных (MUFA), стеариновой (С18:0) и олеиновой (С18:1) жирных кислот были получены наименьшие коэффициенты наследуемости – 0,13-0,19. Максимальные коэффициенты были получены для миристиновой (С14:0) – 0,58, пальмитиновой (С16:0) – 0,41, среднецепочечных (MCFA) – 0,47. Полученные нами результаты согласуются с рядом авторов, также описавших высокую степень наследуемости – свыше 0,4 для С14:0 и С16:0 , , что может быть связано с тем, что молочный жир до 65% представлен насыщенными жирными кислотами, в частности, миристиновой и пальмитиновой, а также путями синтеза жирных кислот. Данные результаты подтверждаются очень высокой генетической корреляцией между жирными кислотами и массовой долей жира: МДЖ%/С14:0 – 0,87, МДЖ%/С16:0 – 0,94, МДЖ%/MCFA – 0,94, МДЖ%/SFA – 0,97. Также очень высокая положительная корреляция была получена для длинноцепочечных жирных кислот (LCFA) и стеариновой (0,89), олеиновой (0,97) жирных кислот; мононенасыщенных жирных кислот и стеариновой (0,78), олеиновой (0,96), длинноцепочечных жирных кислот (0,93). Для полиненасыщенных и транс-изомеров жирных кислот, была также получена очень высокая положительная корреляция (0,77), т.к. TFA являются побочным продуктом образования насыщенных жирных кислот путем гидрогенизации жирных кислот в рубце животного. Образование транс-изомеров жирных кислот из полиненасыщенных и то, что PUFA представлены в основном длинноцепочечными жирными кислотами подтверждается высокой положительной корреляцией между LCFA (0,67), С18:0 (0,61), С18:1 (0,69) и PUFA, а также средней положительной с TFA (0,38/0,46/0,34), низкой корреляцией между TFA/MUFA – 0,26, отрицательной корреляцией между PUFA и C14:0 (-0,19), C16:0 (-0.02), MCFA (-0,06) и средней отрицательной между TFA/C14:0 (-0,41), TFA/C16:0 (-0.33), TFA/MCFA (-0,37). Также низкая отрицательная корреляция между TFA и содержанием жира в молоке и остальными группами жирных кислот может подтверждать влияние трансизомеров ЖК на экспрессию генов, ассоциированных с синтезом молочного жира .
Наследуемость h2 (по диагонали) и генетическая корреляция (под диагональю) между показателями продуктивности и жирнокислотным составом молока
| Сут. удой | МДЖ% | МДБ% | Cas B | C14:0 | C16:0 | C18:0 | C18:1 | LCFA | MCFA | MUFA | PUFA | SFA | SCFA | TFA |
Суточный удой | 0,39 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
МДЖ% | -0,42 | 0,31 |
|
|
|
|
|
|
|
|
|
|
|
|
|
МДБ% | -0,47 | 0,63 | 0,41 |
|
|
|
|
|
|
|
|
|
|
|
|
Cas B | -0,48 | 0,68 | 0,99 | 0,42 |
|
|
|
|
|
|
|
|
|
|
|
C14:0 | -0,46 | 0,87 | 0,61 | 0,68 | 0,58 |
|
|
|
|
|
|
|
|
|
|
C16:0 | -0,42 | 0,94 | 0,56 | 0,60 | 0,89 | 0,41 |
|
|
|
|
|
|
|
|
|
C18:0 | -0,05 | 0,53 | 0,18 | 0,16 | 0,11 | 0,41 | 0,19 |
|
|
|
|
|
|
|
|
C18:1 | -0,21 | 0,66 | 0,43 | 0,41 | 0,25 | 0,48 | 0,83 | 0,13 |
|
|
|
|
|
|
|
LCFA | -0,15 | 0,63 | 0,32 | 0,32 | 0,21 | 0,44 | 0,89 | 0,97 | 0,13 |
|
|
|
|
|
|
MCFA | -0,45 | 0,94 | 0,64 | 0,69 | 0,95 | 0,98 | 0,32 | 0,43 | 0,38 | 0,47 |
|
|
|
|
|
MUFA | -0,30 | 0,77 | 0,47 | 0,48 | 0,43 | 0,59 | 0,78 | 0,96 | 0,93 | 0,56 | 0,13 |
|
|
|
|
PUFA | 0,02 | 0,18 | 0,29 | 0,25 | -0,19 | -0,02 | 0,61 | 0,69 | 0,67 | -0,06 | 0,57 | 0,22 |
|
|
|
SFA | -0,37 | 0,97 | 0,62 | 0,66 | 0,82 | 0,96 | 0,56 | 0,64 | 0,61 | 0,95 | 0,71 | 0,20 | 0,33 |
|
|
SCFA | -0,27 | 0,91 | 0,62 | 0,65 | 0,72 | 0,85 | 0,58 | 0,66 | 0,64 | 0,84 | 0,68 | 0,37 | 0,95 | 0,30 |
|
TFA | 0,15 | -0,14 | -0,05 | -0,09 | -0,41 | -0,33 | 0,46 | 0,34 | 0,38 | -0,37 | 0,26 | 0,77 | -0,16 | -0,03 | 0,24 |
Влияние продолжительности лактации, выраженное через коэффициент регрессии измеряемого признака на количество дойных дней, представлено в таблице 3. Количество дойных дней варьировало в диапазоне от 10 до 1184. Полученные результаты соответствуют изменениям в продуктивности животного в течение лактации, так коэффициент регрессии для удоя составил -0,028, что говорит о естественном уменьшении продуктивности к концу лактации. Напротив, максимальные значения были получены для массовой доли жира и белка и, соответственно, содержания казеинов, а также для среднецепочечных и насыщенных жирных кислот, как максимально представленных в молочном жире.
Коэффициент регрессии показателя количество дойных дней (DIM)
Показатель | Коэффициент регресии |
Суточный удой | -0,028±0,0004 |
МДЖ% | 0,002±0,00007 |
МДБ% | 0,002±0,00002 |
Казеин | 0,002±0,00002 |
Миристиновая (С14:0) | 0,0003±0,000006 |
Пальмитиновая (С16:0) | 0,0005±0,00002 |
Стеариновая (С18:0) | -0,00002±0,00001 |
Олеиновая (С18:1) | 0,0003±0,00002 |
Длинноцепочечные (LCFA) | 0,0002±0,00003 |
Среднецепочечные (MCFA) | 0,001±0,00003 |
Мононенасыщенные (MUFA) | 0,0003±0,00002 |
Полиненасыщенные (PUFA) | 0,00003±0,000002 |
Насыщенные (SFA) | 0,001±0,00004 |
Короткоцепочечные (SCFA) | 0,0002±0,00001 |
Трансизомеры (TFA) | -0,00002±0,000003 |
Стоит отметить, что в ранний период лактации у молочных коров часто наблюдается отрицательный энергетический баланс, что компенсируется активацией резервов организма, а именно запасов собственной жировой ткани, что отражается в профиле жирных кислот и сниженным содержанием коротко- и среднецепочечных жирных кислот , .
Генетические оценки состава молочного жира в зависимости от сезона лактации приведены в таблице 4. Было установлено наибольшее отрицательное влияние летнего периода на продуктивные качества и состав молочного жира коров. Так же отрицательные показатели были получены для насыщенных (SFA), среднецепочечных (MCFA), короткоцепочечных (SCFA) и пальмитиновой (С16:0) с миристиновой (С14:0) жирных кислот в весенний период. Для тех же групп и индивидуальных жирных кислот, а также массовой доли жира и белка было показано максимальное положительное влияние в зимний период. Также зимний период лактации положительно влиял на содержание стеариновой (С18:0), длиноцепочечных (LCFA) и ненасыщенных жирных кислот, но наибольшие значения были показаны для весеннего периода. Различий во влиянии зимнего и весеннего периодов на содержание олеиновой (С18:1) жирной кислоты обнаружено не было. Для транс-изомеров (TFA) жирных кислот положительное влияние было выявлено только в весенний период лактации, также как и для суточного удоя. Данные о снижении содержания насыщенных коротко- и среднецепочечных жирных кислот в летний период и рост в зимний полностью согласуются с литературными данными, но отличные для насыщенных и ненасыщенных длинноцепочечных жирных кислот, где был показан их рост именно в летний период. Данная закономерность была объяснена авторами, как влияние наличия свежескошенной травы в рационе питания, в которой преобладают ненасыщенные жирные кислоты или выпас животных .
Влияние сезона лактации на показатели продуктивности и жирнокислотный состав молока
Показатель | Весна | Зима | Лето | Осень |
Суточный удой | 0,666±0,138 | -1,162±0,152 | -1,218±0,15 | 0,0 |
МДЖ% | -0,099±0,024 | 0,18±0,026 | -0,679±0,026 | 0,0 |
МДБ% | -0,088±0,008 | -0,013±0,009 | -0,218±0,009 | 0,0 |
Казеин | -0,084±0,007 | -0,01±0,008 | -0,223±0,008 | 0,0 |
Миристиновая (С14:0) | -0,031±0,002 | 0,017±0,002 | -0,068±0,002 | 0,0 |
Пальмитиновая (С16:0) | -0,048±0,006 | 0,059±0,006 | -0,139±0,006 | 0,0 |
Стеариновая (С18:0) | 0,015±0,003 | 0,009±0,003 | -0,046±0,003 | 0,0 |
Олеиновая (С18:1) | 0,03±0,008 | 0,03±0,009 | -0,134±0,009 | 0,0 |
Длинноцепочечные (LCFA) | 0,046±0,011 | 0,036±0,012 | -0,187±0,012 | 0,0 |
Среднецепочечные (MCFA) | -0,096±0,009 | 0,087±0,009 | -0,260±0,009 | 0,0 |
Мононенасыщенные (MUFA) | 0,032±0,008 | 0,029±0,009 | -0,106±0,009 | 0,0 |
Полиненасыщенные (PUFA) | 0,007±0,001 | 0,001±0,001 | -0,014±0,001 | 0,0 |
Насыщенные (SFA) | -0,122±0,015 | 0,152±0,017 | -0,519±0,017 | 0,0 |
Короткоцепочечные (SCFA) | -0,027±0,003 | 0,032±0,004 | -0,131±0,004 | 0,0 |
Трансизомеры (TFA) | 0,017±0,001 | -0,001±0,001 | -0,002±0,001 | 0,0 |
Результаты оценки влияния номера лактации на показатели продуктивности и состав жирных кислот демонстрируют общую закономерность в изменении обильномолочности и жирномолочности в зависимости от номера лактации (таблица 5).
Влияние номера лактации на показатели продуктивности и жирнокислотный состав молока
Номер лактации | 1 | 2 | 3 | 4 | 5 |
Суточный удой | 2,632±0,578 | 2,983±0,57 | 1,880±0,58 | 0,983±0,644 | 0,0 |
МДЖ% | -0,69±0,086 | -0,6±0,084 | -0,45±0,087 | -0,092±0,099 | 0,0 |
МДБ% | -0,13±0,035 | -0,10±0,034 | -0,03±0,035 | -0,034±0,038 | 0,0 |
Казеин | -0,12±0,032 | -0,09±0,031 | -0,02±0,032 | -0,017±0,035 | 0,0 |
Миристиновая (С14:0) | -0,15±0,012 | -0,11±0,012 | -0,07±0,012 | -0,018±0,013 | 0,0 |
Пальмитиновая (С16:0) | -0,19±0,025 | -0,14±0,024 | -0,12±0,025 | 0,005±0,027 | 0,0 |
Стеариновая (С18:0) | 0,004±0,008 | -0,004±0,01 | -0,02±0,008 | -0,003±0,010 | 0,0 |
Олеиновая (С18:1) | -0,008±0,020 | -0,04±0,020 | -0,07±0,021 | -0,03±0,026 | 0,0 |
Длинноцепочечные (LCFA) | 0,04±0,027 | -0,02±0,027 | -0,07±0,028 | -0,03±0,035 | 0,0 |
Среднецепочечные (MCFA) | -0,4±0,041 | -0,28±0,040 | -0,21±0,041 | -0,023±0,045 | 0,0 |
Мононенасыщенные (MUFA) | -0,06±0,019 | -0,09±0,019 | -0,09±0,020 | -0,035±0,025 | 0,0 |
Полиненасыщенные (PUFA) | 0,017±0,002 | 0,012±0,002 | 0,01±0,002 | 0,002±0,003 | 0,0 |
Насыщенные (SFA) | -0,31±0,058 | -0,22±0,057 | -0,19±0,059 | 0,021±0,066 | 0,0 |
Короткоцепочечные (SCFA) | -0,03±0,012 | -0,02±0,012 | -0,02±0,012 | 0,011±0,014 | 0,0 |
Трансизомеры (TFA) | 0,013±0,003 | 0,008±0,003 | 0,004±0,003 | -0,001±0,004 | 0,0 |
Показано, что оценка массовой доли жира, а также насыщенных среднецепочечных жирных кислот, увеличивается с каждой последующей лактацией, достигая максимума в 4-ую (пальмитиновая, насыщенные жирные кислоты) или 5-ую лактацию (миристиновая, среднецепочечные жирные кислоты). Влияние на количество длинноцепочечных жирных кислот, существенно отличалось от среднецепочечных, так, максимальная оценка влияния номера лактации продемонстрирована для 1-ой лактации и к 3-ей достигает минимума. Максимально приближенные результаты были показаны и у мононенасыщенных жирных кислот, с той разницей, что максимально отрицательные оценки были получены для 2-ой и 3-ей лактации. Особый интерес представляют полиненасыщенные и транс-изомеры жирных кислот, т.к. для них показаны практически одинаковые тенденции и изменения оценок в зависимости от лактации.
Данные показатели и взаимодействия полностью согласуются с выше полученными данными, особенно в контексте объяснения путей получения жирных кислот молочного жира. Так, для первотелок характерен жирнокислотный профиль с преобладанием ненасыщеных жирных кислот. Авторы это связывают с тем, что первотелки все еще развиваются по сравнению со взрослыми коровами, также была отмечена сниженная экспрессия комплекса ферментов синтетаз жирных кислот в молочной железе первотелок, которая выходит на уровень, характерный для коров последующих лактаций, только к последней трети первой лактации. Таким образом, происходит активное использование жирных кислот из кормов и собственных резервов , , .
Влияние аллельного полиморфизма гена DGAT1 на молочную продуктивность и жирнокислотный состав молока представлено в таблице 6.
Оценки влияния генотипов SNP rs109234250, rs109326954 гена DGAT1 на показатели молочной продуктивности и профиль жирных кислот
Показатель | DGAT1 | ||
AA | AK | KK | |
Суточный удой | -1,674±1,089 | -0,658±0,795 | 0,0 |
МДЖ% | 0,54±0,136 | 0,326±0,099 | 0,0 |
МДБ% | 0,234±0,064 | 0,12±0,047 | 0,0 |
Казеин | 0,215±0,054 | 0,117±0,042 | 0,0 |
Миристиновая (С14:0) | 0,052±0,017 | 0,036±0,012 | 0,0 |
Пальмитиновая (С16:0) | 0,151±0,038 | 0,092±0,028 | 0,0 |
Стеариновая (С18:0) | 0,033±0,012 | 0,017±0,009 | 0,0 |
Олеиновая (С18:1) | 0,125±0,03 | 0,064±0,022 | 0,0 |
Длинноцепочечные (LCFA) | 0,148±0,041 | 0,083±0,03 | 0,0 |
Среднецепочечные (MCFA) | 0,237±0,06 | 0,148±0,044 | 0,0 |
Мононенасыщенные (MUFA) | 0,107±0,029 | 0,058±0,021 | 0,0 |
Полиненасыщенные (PUFA) | 0,011±0,003 | 0,006±0,002 | 0,0 |
Насыщенные (SFA) | 0,406±0,092 | 0,243±0,067 | 0,0 |
Короткоцепочечные (SCFA) | 0,09±0,02 | 0,054±0,014 | 0,0 |
Транс-изомеры (TFA) | 0,004±0,005 | 0,003±0,003 | 0,0 |
Влияние полиморфизма rs109234250, rs109326954 гена DGAT1 на молочную продуктивность и состав жирных кислот показало положительную ассоциацию для всех рассматриваемых показателей для аллеля А, кроме удоя, где желательным аллелем является аллель К. Полученные результаты отличались от имеющихся данных, так для разводимых в нашей стране пород скота в основном было показано положительное влияние аллеля А на обильномолочность, в то время как на жирномолочность больше влиял аллель К , , , но обратный результат был получен для голштинизированных симменталов . Среди иностранных исследователей, также имеются противоречивые результаты, вплоть до отсутствия достоверных ассоциаций. Однозначного влияние какого-либо из аллеля гена DGAT1 на жирнокислотный состав молочного жира также не представлено. Так, положительное влияние аллеля А, показанное E.Samková et al. 2021 для насыщенных коротко- и среднецепочечных жирных кислот и положительное влияние аллеля К на ненасыщенные жирные кислоты, не согласуются с ранее полученными данными A.M. Carvajal et al 2015 и Schennink et al. 2008 , Ассоциация аллеля К с длинноцепочечными жирными кислотами, у последних авторов, отличалась от результатов G. Conte et al. 2010 , который показал взаимосвязь с аллелем А.
Влияние аллеля А на содержание транс-изомеров жирных кислот хоть и было показано, но достоверных отличий обнаружено не было.
4. Заключение
В нашем исследовании было изучено влияние на молочную продуктивность и состав жирных кислот таких факторов, как возраст животного, выраженный в номере лактации, продолжительность и сезон лактации, а также полиморфизм гена DGAT1. Полученные результаты предполагают возможность изменения профиля жирных кислот молочного жира в результате отбора и селекции. Но стоит учитывать, что селекционная работа в равной степени может регулироваться скорректированным рационом кормления с содержанием желательных жирных кислот. Использование информации о генотипах гена DGAT1 в полной мере на данный момент затруднительно, т.к. наибольшее влияние аллельного полиморфизма показано для содержания жира в молоке, что, возможно, корректирует оценки жирнокислотного состава, но однозначных сведений о влиянии определенного аллеля нет. Это свидетельствует о необходимости дополнительных дальнейших исследований в данном направлении с расширением исследовательской базы и параметров.