Polymorphism of the KAP1.1 gene in domestic sheep
Polymorphism of the KAP1.1 gene in domestic sheep

Abstract
Keratin proteins are a key factor determining wool production in animals. Variation in the KRTAP1.1 gene, which is responsible for the synthesis of the KAP1.1 protein, potentially influences the wool quality of sheep. The study assessed the distribution of polymorphism in this gene across 10 populations of domestic sheep breeds. The results showed a predominance of the B allele and the BB genotype within the examined populations. The C allele was present to a lesser extent, whilst the A allele was virtually absent in the tested animals. The greatest genetic diversity at this locus was observed in animals of the Romanovsk, Groznensk and Caucasian sheep breeds, and the least in the Kuibyshevsk breed. In terms of the distribution of the studied polymorphism, the Romanovsk and Edilbaevsk breed populations differed significantly from the other breeds.
1. Введение
Овцеводство на протяжении веков является одной из важнейших отраслей животноводства России и служит источником таких незаменимых продуктов, как мясо, шерсть и молоко. Шерсть овец используется для производства широкого спектра текстильной продукции, начиная от простых валяных изделий и заканчивая качественными дорогостоящими костюмами. Несмотря на повсеместный переход легкой промышленности на синтетическое сырье, ткани из натуральных волокон и овечья шерсть в особенности остаются востребованы благодаря их гигроскопичности, эластичности, мягкости, стойкости к накоплению запахов, способности держать тепло, при этом сохраняя вентилирующие качества, а также меньшим влиянием на экологию при производстве и переработке в сравнении с искусственными волокнами , .
Получение шерсти высокого качества требует использования современных методов селекции овец. Маркерная селекция (MAS — marker assisted selection), является одним из таких методов. В рамках данного метода проводится молекулярно-генетическая диагностика с целью выявления генетических маркеров тех или иных хозяйственно-полезных признаков животных.
Среди генов, контролирующих формирование шерстного волокна, выделяют KAP (keratin associated proteins), отвечающие за синтез белков матрикса, окружающего волокна кератина и KRT (keratins), непосредственно гены кератиновых волокон. Существует целый ряд исследований, которые показывают связь данных генов с тониной, изменчивостью тонины, длиной и прочностью штапеля, величиной настрига и другими качествами, учет которых важен при производстве текстильной промышленности .
Одним из маркеров, потенциально влияющих на шерстные качества овец, является KAP1.1. У овец ген KRTAP1.1 (ранее известный как B2A) расположен на 11 хромосоме . В норме KRTAP1.1 содержит четыре декапептидных последовательности вида QPTSIQTSCC, первоначально обозначенный Rogers G.R. с соавторами как QTSCCQPTSI , . Изменчивость данного гена характеризуется появлением двух возможных мутаций — одной инсерции и одной делеции длиной по 30 пар нуклеотидов, кодирующих вышеупомянутую аминокислотную последовательность, что приводит к возникновению трех аллельных вариаций KRTAP1.1 . Наличие этих мутаций у овец различных пород было отмечено в ряде работ. Так, в исследовании Itenge-Mweza T.O. с соавторами была отмечена полиморфность данного гена в популяции мериносов Новой Зеландии, и были выявлены все три аллельные вариации . В последующих работах этого же автора была показана достоверная связь этого полиморфизма с показателями выхода мытой шерсти и длиной штапеля , . В работе Ibrahim M.F. с соавторами был отмечен полиморфизм данного гена у египетских пород овец и его связь с показателями изменчивости тонины, настрига мытой шерсти, длины и прочности штапеля .
Цель данной работы — выявить полиморфизм по гену KAP1.1 у отечественных пород овец и оценить его распределение в структуре популяций.
2. Методы и принципы исследования
Для анализа были взяты пробы крови овец пород куйбышевская (популяции Кб-1 и Кб-2), дагестанская горная (популяции Дг-1 и Дг-2), романовская (популяции Рм-1 и Рм-2), кавказская, эдильбаевская, грозненская и черноземельский меринос. Из крови получали лейкоциты с последующим выделением ДНК с помощью наборов ДНК-Экстран 2 (Синтол, г. Москва). Полученный в результате ПЦР амплификат был проанализирован с использованием системы гель-документирования GelDoc XR+ (BioRad, США). Олигонуклеотиды для постановки ПЦР были произведены компанией Евроген (г. Москва). Последовательность праймеров согласно работе Itenge-Mweza T.O. с соавторами :
F: 5-CAA CCC TCC TCT CAA CCC AAC TCC-3 R: 5-GCT GCT ACC CAC CTG GCC ATA-3.
Программа амплификации:
95°C — 2 мин., 40 циклов (95°C — 20 сек., 62°C — 30 сек., 72°C — 30 сек.), 72°C — 5 мин.
Для проведения расчетов и создания рисунков были использованы программы: MS Excel, PopGen32, Genepop, MEGA и SPSS.
3. Основные результаты
В результате анализа были выявлены аллели A, B и C (ампликоны длиной 341, 311 и 281 п.н. соответственно) и их возможные сочетания: генотипы AA, AB, AC, BB, BC и CC. Результаты электрофоретического разделения полученных фрагментов представлены на рисунке 1. На рисунке 2 изображены гистограммы показателей индекса Шеннона и эффективного числа аллелей. Дендрограмма на основе генетического расстояния между популяциями изображена на рисунке 3. Частоты выявленных аллелей и генотипов представлены в таблице 1. В таблице 2 представлены популяционно-генетические показатели изучаемых популяций. Матрица с показателями уровня значимости (p) по результатам точного G-теста представлена в таблице 3. Показатели генетического сходства и расстояния представлены в таблице 4.
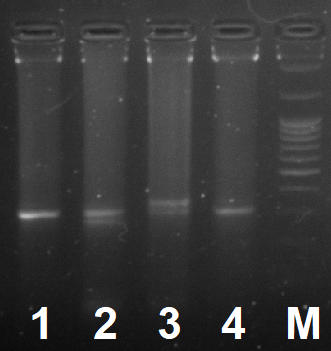
Пример полиморфизма длин полученных фрагментов в агарозном геле (3%)
1 и 4 дорожки содержат генотип BB, а 2 и 3 – генотипы BC и AB соответственно; M – дорожка маркера молекулярного веса (100 bp+1.5 Kb+3 Kb)
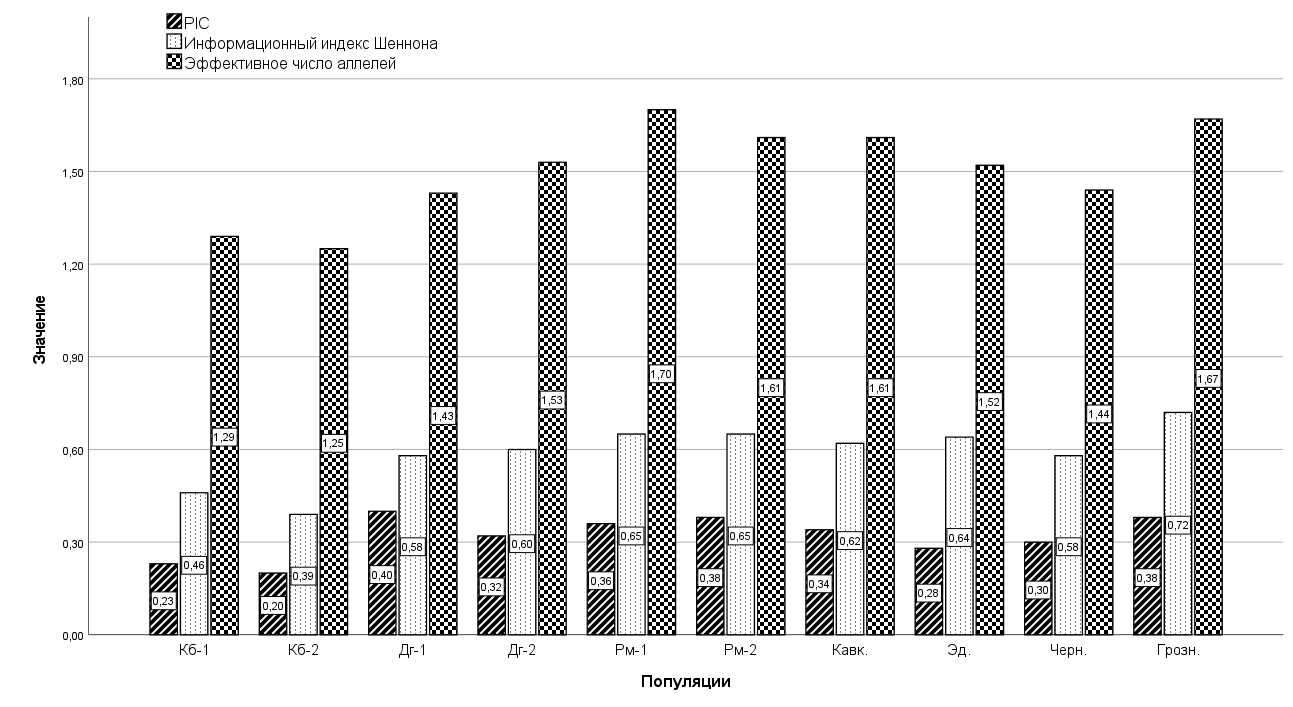
Гистограмма по показателям PIC, индекса Шеннона и эффективного числа аллелей
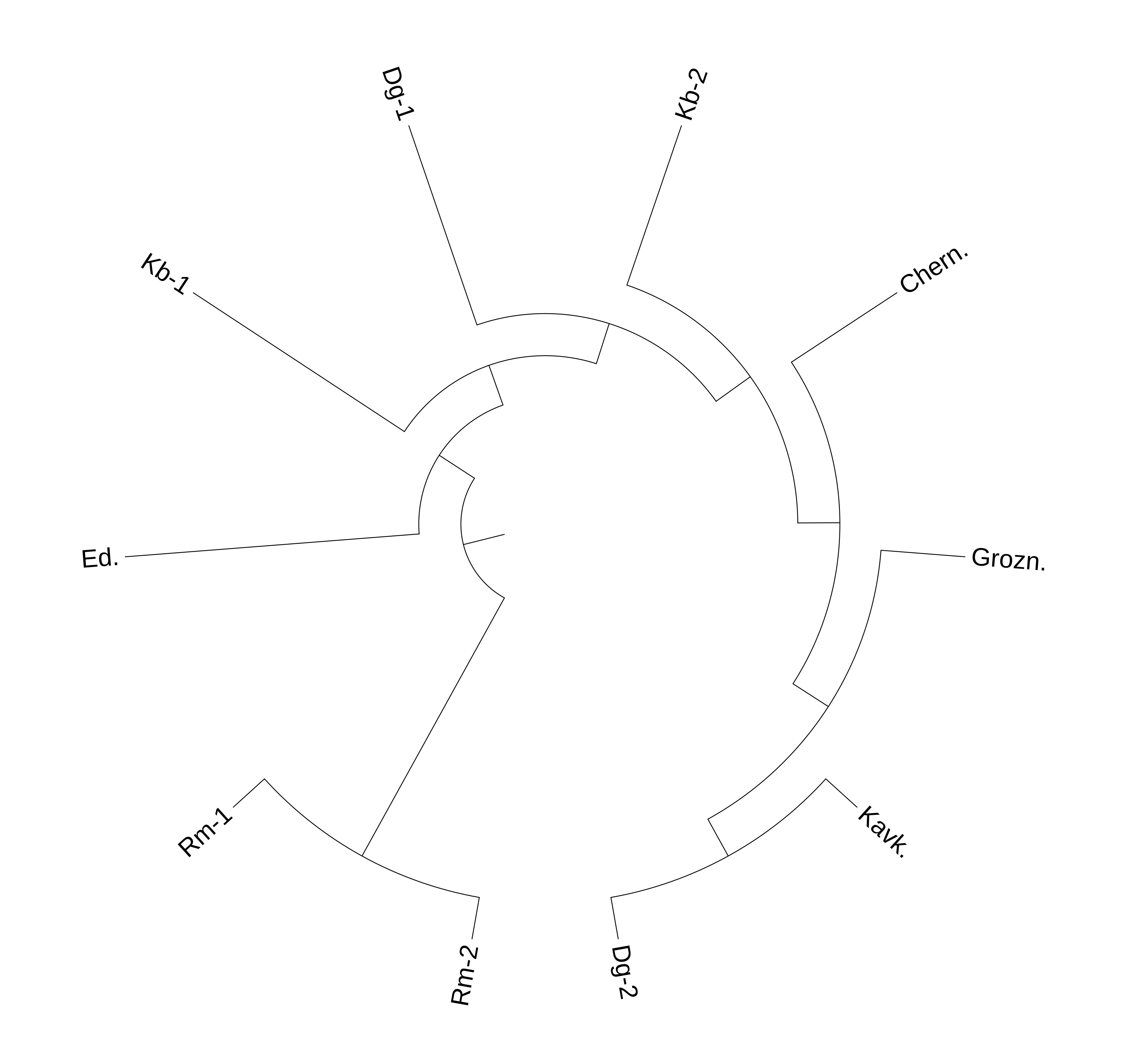
Круговая дендрограмма на основе генетического расстояния между популяциями
Таблица 1 - Распределение частот аллелей и генотипов гена KAP1.1 по 10 популяциям овец
Популяция | Частоты аллелей | Частоты генотипов | |||||||
A | B | C | AA | AB | AC | BB | BC | CC | |
Кб-1 (n=72) | 0,05 | 0,87 | 0,08 | 0,02 | 0,07 | 0 | 0,76 | 0,15 | 0 |
Кб-2 (n=68) | 0,01 | 0,89 | 0,1 | 0 | 0,03 | 0 | 0,79 | 0,16 | 0,02 |
Дг-1 (n=40) | 0,06 | 0,83 | 0,11 | 0,025 | 0,075 | 0 | 0,675 | 0,225 | 0 |
Дг-2 (n=35) | 0,03 | 0,78 | 0,19 | 0 | 0,057 | 0 | 0,629 | 0,257 | 0,057 |
Рм-1 (n=34) | 0,27 | 0,72 | 0,01 | 0,06 | 0,38 | 0,03 | 0,53 | 0 | 0 |
Рм-2 (n=40) | 0,2 | 0,76 | 0,04 | 0,025 | 0,35 | 0 | 0,55 | 0,075 | 0 |
Кавказская (n=63) | 0,02 | 0,75 | 0,23 | 0 | 0,03 | 0 | 0,62 | 0,24 | 0,11 |
Эдильбаевская (n=59) | 0,13 | 0,8 | 0,07 | 0,02 | 0,19 | 0,03 | 0,64 | 0,12 | 0 |
Черноземельская (n=42) | 0,06 | 0,82 | 0,12 | 0 | 0,12 | 0 | 0,67 | 0,19 | 0,02 |
Грозненская (n=36) | 0,08 | 0,75 | 0,17 | 0 | 0,16 | 0 | 0,53 | 0,28 | 0,03 |
Таблица 2 - Популяционно-генетические показатели
Популяция | Ho1 | He2 | Fis3 | ne4 | I5 | PIC6 | HWE7 |
Кб-1 | 0,22 | 0,23 | 0,02 | 1,29 | 0,46 | 0,23 | 0,26 |
Кб-2 | 0,19 | 0,2 | 0,04 | 1,25 | 0,39 | 0,20 | 0,58 |
Дг-1 | 0,3 | 0,31 | 0,01 | 1,43 | 0,58 | 0,28 | 0,21 |
Дг-2 | 0,31 | 0,35 | 0,1 | 1,53 | 0,6 | 0,34 | 0,73 |
Рм-1 | 0,41 | 0,42 | -0,003 | 1,7 | 0,65 | 0,40 | 0,41 |
Рм-2 | 0,43 | 0,38 | -0,13 | 1,61 | 0,65 | 0,36 | 0,76 |
Кавказская | 0,27 | 0,38 | 0,29 | 1,61 | 0,62 | 0,38 | 0,047 |
Эдильбаевская | 0,34 | 0,35 | 0,01 | 1,52 | 0,64 | 0,32 | 0,71 |
Черноземельская | 0,31 | 0,311 | -0,01 | 1,44 | 0,58 | 0,30 | 0,59 |
Грозненская | 0,44 | 0,41 | -0,1 | 1,67 | 0,72 | 0,38 | 0,74 |
Примечание: 1 – наблюдаемая гетерозиготность; 2 – ожидаемая гетерозиготность; 3 – индекс фиксации; 4 – эффективное число аллелей; 5 – информационный индекс Шеннона; 6 – показатель информативности полиморфизма; 7 – показатель уровня значимости (p) отклонения распределения генотипов от равновесного по закону Харди-Вайнберга
Таблица 3 - Показатели уровня значимости (p) по результатам точного G-теста
Популяция | Кб-1 | Кб-2 | Дг-1 | Дг-2 | Рм-1 | Рм-2 | Кавк. | Эд. | Черн. | Грозн. |
Кб-1 | 1 | 0,29 | 0,65 | 0,1 | 1,60e-05 | 0,002 | 0,004 | 0,09 | 0,55 | 0,1 |
Кб-2 | 0,29 | 1 | 0,20 | 0,2 | <2,00e-06 | 6,00e-06 | 0,019 | 0,0011 | 0,19 | 0,02 |
Дг-1 | 0,65 | 0,20 | 1 | 0,39 | 0,0007 | 0,012 | 0,071 | 0,28 | 1 | 0,57 |
Дг-2 | 0,1 | 0,2 | 0,39 | 1 | <2,00e-06 | 0,0003 | 0,66 | 0,01 | 0,43 | 0,42 |
Рм-1 | 1,60e-05 | <2,00e-06 | 0,0007 | <2,00e-06 | 1 | 0,51 | <2,00e-06 | 0,02 | 0,0004 | 0,0002 |
Рм-2 | 0,002 | 6,00e-06 | 0,012 | 0,0003 | 0,51 | 1 | <2,00e-06 | 0,25 | 0,008 | 0,006 |
Кавк, | 0,004 | 0,019 | 0,071 | 0,66 | <2,00e-06 | <2,00e-06 | 1 | 5,80e-05 | 0,07 | 0,08 |
Эд, | 0,09 | 0,0011 | 0,28 | 0,01 | 0,02 | 0,25 | 5,80e-05 | 1 | 0,19 | 0,13 |
Черн, | 0,55 | 0,19 | 1 | 0,43 | 0,0004 | 0,008 | 0,07 | 0,19 | 1 | 0,57 |
Грозн, | 0,1 | 0,02 | 0,57 | 0,42 | 0,0002 | 0,006 | 0,08 | 0,13 | 0,57 | 1 |
Таблица 4 - Показатели генетического сходства (сверху от диагонали) и расстояния (снизу от диагонали)
Поп-я | Кб-1 | Кб-2 | Дг-1 | Дг-2 | Рм-1 | Рм-2 | Кавк. | Эд. | Черн. | Грозн. |
Кб-1 | - | 0,9990 | 0,9986 | 0,9893 | 0,9542 | 0,9792 | 0,9776 | 0,9947 | 0,9983 | 0,9900 |
Кб-2 | 0,0010 | - | 0,9979 | 0,9920 | 0,9407 | 0,9698 | 0,9821 | 0,9900 | 0,9978 | 0,9896 |
Дг-1 | 0,0014 | 0,0021 | - | 0,9946 | 0,9557 | 0,9800 | 0,9856 | 0,9958 | 1,0000 | 0,9960 |
Дг-2 | 0,0107 | 0,0080 | 0,0054 | - | 0,9294 | 0,9596 | 0,9978 | 0,9833 | 0,9955 | 0,9973 |
Рм-1 | 0,0469 | 0,0611 | 0,0453 | 0,0733 | - | 0,9950 | 0,9100 | 0,9785 | 0,9538 | 0,9521 |
Рм-2 | 0,0210 | 0,0307 | 0,0202 | 0,0413 | 0,0050 | - | 0,9429 | 0,9941 | 0,9787 | 0,9753 |
Кавк. | 0,0226 | 0,0180 | 0,0145 | 0,0022 | 0,0944 | 0,0588 | - | 0,9707 | 0,9871 | 0,9931 |
Эд. | 0,0053 | 0,0101 | 0,0042 | 0,0168 | 0,0217 | 0,0059 | 0,0297 | - | 0,9913 | 0,9952 |
Черн. | 0,0017 | 0,0022 | 0,0000 | 0,0045 | 0,0473 | 0,0216 | 0,0130 | 0,0049 | - | 0,9966 |
Грозн. | 0,0100 | 0,0104 | 0,0040 | 0,0027 | 0,0491 | 0,0251 | 0,0069 | 0,0087 | 0,0034 | - |
4. Обсуждение
Как и в работах других исследователей, в структуре изучаемых популяций преобладал аллель B и генотип BB. Наибольшая их частота была замечена у куйбышевской породы, составляя 0,89 по содержанию аллеля B и 0,79 по содержанию генотипа BB в структуре популяции Кб-2. Наименьшая частота аллея B и генотипа BB наблюдалась у животных романовской (0,72 по аллелю B и 0,53 по генотипу BB у Рм-1) и грозненской (0,75 по аллелю B и 0,53 по генотипу BB) породы овец.
Гораздо более редкими оказались аллели A и C и содержащие их генотипы. Так, самое низкое содержание аллеля A наблюдалось у животных куйбышевской породы (0,01 у Кб-2), а самое высокое — у животных романовской породы (0,27 у Рм-1). Гомозиготный генотип AA отсутствовал у половины популяций, его наибольшая частота была отмечена у животных популяции Рм-1. Гетерозиготный генотип AB чаще всего встречался у животных романовской породы (0,38 у Рм-1), а его наименьшая частота была отмечена у кавказской породы (0,03) и куйбышевской породы (0,03 у Кб-2). Аллель C был представлен в наибольшей степени у животных кавказской породы (0,23) и в наименьшей степени у животных романовской породы (0,01 у Рм-1). Гомозиготный генотип CC, как и в случае с AA, присутствовал лишь у половины популяций, а его наибольшая частота составила 0,11 у животных кавказской породы. Гетерозиготный генотип BC в наибольшей степени был представлен у животных грозненской породы (0,28), а в наименьшей — у романовской породы, где у популяции Рм-2 его частота составила 0,075 (у Рм-1 генотип BC отсутствовал). Наиболее редкий генотип AC отсутствовал у большей части популяций и был обнаружен только у животных романовской и эдильбаевской пород (0,03 у обеих популяций).
При сравнении полученных данных с результатами зарубежных исследователей стоит отметить заметное преобладание аллеля B и генотипа BB в структуре популяций отечественных пород. Так, в турецком исследовании Yardibi H. с соавторами содержание аллеля B и генотипа BB в структуре популяций было заметно ниже, а наибольшее их содержание было отмечено у породы Kivircik (0,7 и 0,6 соответственно) . В работе Itenge-Mweza T.O. с соавторами от 2019 года частоты аллеля B и генотипа BB у африканской породы Swakara составили 0,57 и 0,35 соответственно . Таким образом, у исследуемых в данной работе животных минимальная частота аллеля B превышает максимальную частоту этого же аллеля у зарубежных пород.
Стоит также отметить, что наибольшее содержание аллеля A и генотипа AA наблюдается у пород овец, имеющих грубую шерсть (романовская, эдильбаевская), в то время как аллель C и генотип CC были больше представлены у пород, характеризующихся тонкой шерстью (дагестанская, черноземельская, кавказская, грозненская).
Все популяции находились в равновесном состоянии, согласно закону Харди-Вайнберга, за исключением кавказской породы овец, у которой показатель значимости точного теста был ниже порогового значения (0,05).
Для более точной оценки аллельных равновесий в популяциях были посчитаны показатели ожидаемой и наблюдаемой гетерозиготности, а также индекс фиксации (Fis), также известный как коэффициент инбридинга. За исключением кавказской и романовской (Рм-2) пород, все популяции имели близкие к нулю значения индекса фиксации, показывая незначительные различия между ожидаемой и наблюдаемой гетерозиготностью. Рм-2 показала повышенную степень аутбридинга (-0,13). Кавказская порода показала высокий показатель индекса фиксации (0,29), что может свидетельствовать о значительной степени инбридинга в данной популяции. Тем не менее, данные по одному локусу и на довольно ограниченном объеме выборки недостаточно репрезентативны для однозначной оценки степени инбридинга по всей отаре.
Для более детальной оценки генетического разнообразия популяций были рассчитаны показатели PIC (информативность полиморфизма), индекса Шеннона (I) и эффективного числа аллелей (ne), по которым была построена гистограмма. При расчете обоих показателей за основу берутся частоты аллелей, но особенности расчета приводят к тому, что показатель I более чувствителен к наличию редких аллелей, в то время как PIC и ne сильно зависят от наличия доминирующих аллелей. В комплексе с наблюдаемой гетерозиготностью эти три показателя позволяют лучше оценить аллельное разнообразие популяции. К наиболее полиморфным популяциям можно отнести животных романовской, грозненской и кавказской пород. Чуть меньшие значения показали популяции эдильбаевской, черноземельской и дагестанской пород. Самым низким разнообразием обладали животные куйбышевской породы.
Для достоверной оценки отличий между популяциями был использован попарный точный G-тест. При заданном уровне значимости (0,0011 с поправкой Бонферрони) наиболее характерными отличиями от остальных популяций обладали животные романовской породы, которые достоверно отличались от большинства популяций.
По результатам оценки генетического расстояния популяции романовской породы были отнесены в отдельный кластер, а также, наряду с эдильбаевской, показали наибольшее отличие от остальных пород по исследуемому полиморфизму. Наибольшее расстояние отмечалось между романовской и кавказской породами овец. Стоит отметить, что характерное отличие романовской и эдильбаевской популяций от остальных исследуемых животных может быть обусловлено сельскохозяйственной направленностью данных пород. Так, и романовская, и эдильбаевская породы имеют грубую шерсть, тогда как для остальных исследуемых пород характерно тонкое и полутонкое руно.
5. Заключение
По итогам данной работы было выявлено, что у отечественных пород преобладет генотип BB и аллель B. В меньшей степени был представлен аллель C и генотипы BC и AB. Аллель A был представлен в наименьшей степени, а генотипы AA, AC и CC были найдены лишь у отдельных животных, либо отсутствовали. При этом достоверное отличие от остальных исследуемых популяций в распределении аллелей и генотипов было отмечено у обеих популяции романовской породы овец и у животных эдильбаевской породы овец, имевших в своей структуре наибольшую встречаемость аллеля A и содержащих его генотипов. Причиной тому, как отмечалось выше, может являться сельскохозяйственная направленность данных пород.
Все популяции, за исключением кавказской породы, находились в равновесном состоянии, согласно закону Харди-Вайнберга. Также у животных кавказской породы был отмечен высокий коэффициент инбридинга. Наиболее полиморфными по исследуемой мутации были животные романовской, грозненской и кавказской пород, в то время как животные куйбышевской породы обладали наименьшим разнообразием. По распределению генотипов и аллелей наиболее отличались от остальных популяций животные романовской породы.
Для более полной оценки данной мутации у отечественных пород рекомендуется увеличить объем исследуемых популяций, а также проверить связь данного полиморфизма с шерстными показатели продуктивности.