ДИНАМИКА ЖИВОГО НАПОЧВЕННОГО ПОКРОВА КУЛЬТУР СОСНЫ И ЕЛИ В ЗАВИСИМОСТИ ОТ УСЛОВИЙ ПРОИЗРАСТАНИЯ
ДИНАМИКА ЖИВОГО НАПОЧВЕННОГО ПОКРОВА КУЛЬТУР СОСНЫ И ЕЛИ В ЗАВИСИМОСТИ ОТ УСЛОВИЙ ПРОИЗРАСТАНИЯ

Аннотация
Структура травянистого яруса лесных ценозов, его количественный и качественный состав во многом определяются микроклиматом среды, который формируется и изменяется по мере роста и развития лесных культур. Данные наблюдений за лесными культурами в разных фазах роста наглядно иллюстрируют динамику происходящей сукцессии растительных сообществ.
Цель исследований – изучение динамики видового разнообразия живого напочвенного покрова лесных культур в зависимости от их породного состава и возраста, а также условий произрастания.
Проведены мониторинговые исследования живого напочвенного покрова лесных культур. В результате исследований пришли к выводу, что после антропогенного нарушения фитоценозов происходит трансформация эколого-ценотической структуры флоры в направлении восстановления исходной структуры лесной среды. Характер изменений происходит по пути постепенного вытеснения луговых видов активной лесной группой растений, а также за счет сокращения доли менее устойчивых неактивных и малоактивных видов.
1. Введение
Изучение растительности нижних ярусов на лесокультурных площадях играет важную роль в исследовании скорости лесовозобновительного процесса. Напочвенному покрову в фитоценозе отведена значимая ценотическая роль: растения способствуют увеличению мощности плодородного горизонта почвы, повышают активность микрофлоры и зоокомпонента почвы и др.
В связи с этим необходимо всестороннее изучение растений напочвенного покрова, определение наиболее активных видов, которые непременно влияют на ход лесовозобновительных процессов.
Применительно к лесохозяйственной практике предлагаем принять за основу активности видов – способность растений участвовать в формировании и смене растительных сообществ с помощью многообразных механизмов адаптации, которые в своей совокупности помогают преодолевать негативное воздействие неблагоприятных факторов, выживать, размножаться и быть устойчивыми в фитоценозах. Классификация активности видов является теоретической основой оценки антропогенной нагрузки в ценозе, а также служит сигналом для выявления значимых видов.
2. Объекты и методы исследований
Тест-объектами наших исследований явились культурфитоценозы антропогенно-природного происхождения сосны (Pinus sylvestris L.) и ели (Picea abies (L.) H. Karst.) в фазе смыкания (14 лет) и чащи (22 года), то есть до формирования исконно лесной среды. Объекты исследований расположены на территории Архангельской области.
В работе использовали общепринятые в лесоводстве и таксации методики исследований древостоя .
Количественный анализ флоры или парциальная активность видов (ПарцА) оценивалась на основании методических подходов Б.А. Юрцева . В основу фитоценотической классификации активности видов растений (ФитоА) по широте эколого-ценотической амплитуды легли геоботанические исследования , , , . На основе проведенного анализа методических подходов к определению активности видов в работе была предложена и использована адаптированная классификация активности растений напочвенного покрова культурфитоценозов в условиях северной части Восточно-Европейской равнины. Усредненная таксационная характеристика исследуемых древостоев представлена в таблице 1.
Таблица 1 - Усредненные таксационные показатели культур
Вариант | Густота, тыс. шт./га | Средний диаметр, см | Средняя высота2, м | Класс бонитета | Полнота | Запас древесины, м3/га | |
первоначальная | на момент исследования | ||||||
Кисличный тип лесорастительных условий Дренированные условия в возрасте1 14 лет | |||||||
Сосна | 2,5 | 2,2 | 8,3 | 6,3 | I | 0,8 | 69,9 |
Ель | 3,5 | 2,9 | – | 1,8 | V | – | – |
22 года | |||||||
Сосна | 2,5 | 2,1 | 12,6 | 11,3 | Iа | 1,1 | 164,7 |
Ель | 3,5 | 1,1 | 6,7 | 5,3 | IV | 0,4 | 18,1 |
Черничный тип лесорастительных условий Условия временно избыточного увлажнения в возрасте1 14 лет | |||||||
Сосна | 2,5 | 1,6 | 6,9 | 5,7 | II | 0,4 | 24,7 |
Ель | 2,8 | 2,4 | – | 1,8 | V | – | – |
22 года | |||||||
Сосна | 2,5 | 1,4 | 11,2 | 9,8 | I | 0,6 | 70,3 |
Ель | 2,8 | 2,1 | 5,1 | 4,5 | IV | 0,4 | 16,1 |
Примечание: 1 - с момента создания культур; 2 – при средней высоте древостоя менее 3 м в качестве ведущего показателя принята высота культур в соответствии с рекомендациями В.В. Огиевского, А.А. Хирова [6]
Ландшафтная активность (ЛандА) определена путем суммированных парциальных оценок с учетом широты эколого-ценотической амплитуды:
где ЛандАi – ландшафтная активность i-го вида (балл), ФитоАi – балл фитоценотической активности i-го вида, ПарцАi – балл парциальной активности i-го вида, Pi – балл постоянства i-го вида, Oi – балл обилия i-го вида, N – число описаний.
3. Основные результаты
При выполнении сравнительного анализа флористических показателей напочвенного покрова, исследованных нами культурфитоценозов, произрастающих в разных условиях местопроизрастания, следует отметить, что для насаждений черничного типа леса в условиях временного избыточного переувлажнения значения показателей видового богатства травянистых растений ниже, чем в кисличном типе леса на дренированных почвах.
Такой характер развития живого напочвенного покрова связан с особенностью временно переувлажненных экосистем, которые имеют более высокую влажность в сравнении с дренированными почвами. Избыточное увлажнение ведет к изменению температурного режима почв, снижению ее аэрации и, как следствие, уменьшению объемной массы и повышению кислотности. Такое сочетание факторов среды ведет к формированию природных условий, в которых может существовать строго ограниченный видовой состав растений.
В кисличном типе леса к моменту смыкания культур сосны и ели в живом напочвенном покрове выявлено 36 и 33 вида растений соответственно, в том числе 2 вида кустарничков (Vaccinium vitis-idaea L., Vaccinium myrtillus L.), 2 вида моховой растительности (Polytrichumcommune Hedw., Pleuroziumschreberi (Willd. Ex Brid.) Mitt.) и 2 малолетних травянистых вида (Melampyrum pratense L., Campanula patula L.) (рис. 1).
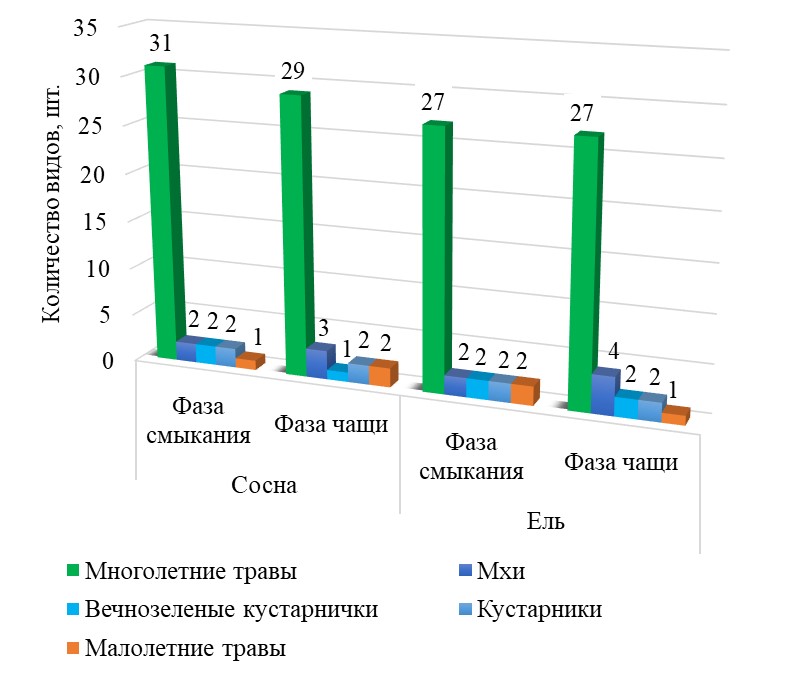
Видовое богатство растительных сообществ культур сосны и ели на дренированных почвах
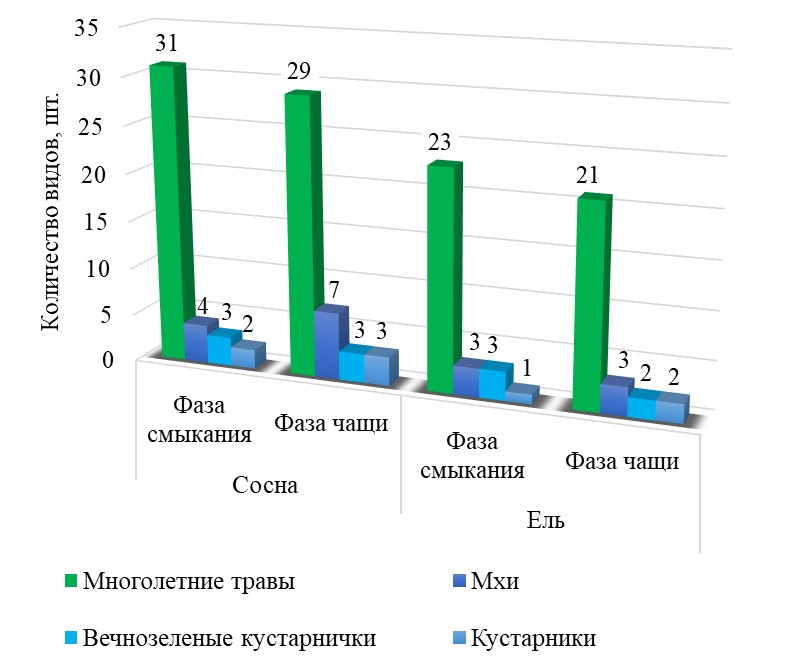
Видовое богатство растительных сообществ культур сосны и ели в условиях периодического переувлажнения почв
Относительное участие видов разнотравья и кустарничков на всех исследуемых участках с момента смыкания древостоя до достижения ими фазы чащи постепенно снижается в диапазоне от 12 до 45%.
Такой зрительный эффект упрощенности структуры сообщества, снижение видового разнообразия при движении к климаксу свойственны таежной зоне . По мнению ряда авторов, внутренняя структура ценопопуляции усложняется только за счет возрастной дифференциации, а структура сообщества в целом – за счет мозаичности , , , .
Проведенный нами анализ доказывает возрастные изменения видового состава растительных сообществ. К фазе чащи происходит постепенное уменьшение общего числа видов, при этом существует достоверная (p = 0,05) высокая связь между общим числом видов и возрастом лесных культур (r = -0,77).
Как показывают наши данные, флористический состав живого напочвенного покрова с возрастом культур существенно видоизменяется. На смену луговым растениям, которые характерны для открытых пространств, приходят типично лесные виды. Изменения в составе луговой растительности особенно заметны на примере дренированных почв, поскольку на данном этапе формирования культур древесный ярус здесь отличается более высокими таксационными параметрами (табл. 1).
К 22-летнему возрасту лесных культур с момента создания мхи увеличились в видовом количественном отношении в 1,8 раза: отмечены новые виды Hylocomium splendens Hedw., Plagiomnium undulatum (Hedw.) T.J. Kop., Ptilium crista-castrensis (Hedw.) De Not., Rhytidiadelphus triquetrus (Hedw.) Warnst.
Как нам кажется, оценка флористического состава недостаточно полно характеризует ход сукцессии растительных сообществ. В этой связи для характеристики богатства и изменчивости живого напочвенного покрова лесных культур мы использовали классификацию активности видов. Она качественно дополняет флористический анализ, тем самым позволяет оценить иерархические позиции видов в растительном сообществе (табл. 2.).
Таблица 2 - Систематическая структура растительных сообществ на участках культур сосны и ели
Семейство | Количество видов в условиях | |||||||
дренированных почв | временного переувлажнения | |||||||
сосна | ель | сосна | ель | |||||
фаза смыкания | фаза чащи | фаза смыкания | фаза чащи | фаза смыкания | фаза чащи | фаза смыкания | фаза чащи | |
Неактивные виды | ||||||||
Бобовые Fabaceae | - | 1 | - | - | 2 | - | - | - |
Вересковые Ericaceaе | 1 | - | 1 | 1 | - | - | - | - |
Губоцветные Lamiaceae | - | - | - | - | - | 1 | - | - |
Злаковые Poaceae | - | - | - | - | - | 1 | 1 | - |
Зонтичные Apiaceae | 2 | - | 1 | - | - | 1 | - | - |
Колокольчиковые Campanulaceae | 1 | 1 | 1 | - | - | - | - | - |
Лилейные Liliaceae | 1 | 1 | - | 1 | - | - | - | - |
Лютиковые Ranunculaceae | - | 1 | 1 | - | - | - | - | - |
Норичниковые Scrophulariaceae | 2 | - | 2 | 1 | - | 1 | - | - |
Первоцветные Primulaceae | - | - | - | - | 1 | - | - | - |
Розовые Rosaceae | 1 | - | 1 | 1 | - | - | - | - |
Сложноцветные Asteracea | 2 | - | 1 | - | 1 | - | 1 | - |
Хвощевые Equisetaceae | 1 | - | 1 | - | 1 | 1 | - | - |
Всего видов | 11 | 4 | 9 | 4 | 5 | 5 | 2 | 0 |
Малоактивные виды | ||||||||
Аспидиевые Aspidiaceae | - | - | - | - | - | 1 | - | - |
Бобовые Fabaceae | - | - | - | - | - | 1 | - | 1 |
Гвоздичные Caryophyllaceae | 1 | 1 | 1 | 1 | - | - | - | - |
Гераниевые Geraniaceae | 1 | - | 1 | - | - | - | - | - |
Жимолостные Capryfoliaceae | - | - | - | - | 1 | 1 | 1 | - |
Злаковые Poaceae | 1 | - | 1 | - | 1 | - | - | - |
Зонтичные Apiaceae | - | - | - | - | 1 | - | - | - |
Кисличные Oxalidaceae | 1 | - | - | - | 1 | - | 1 | - |
Кочедыжниковые Athyriaceae | - | - | - | - | 1 | - | 1 | - |
Крапивные Urticaceae | 1 | - | 1 | - | - | - | - | - |
Лилейные Liliaceae | - | - | - | - | 1 | - | - | 1 |
Мареновые Rubiaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Норичниковые Scrophulariaceae | - | 1 | - | 1 | - | - | - | - |
Осоковые Cyperaceae | - | - | - | - | 1 | - | 1 | - |
Первоцветные Primulaceae | - | 1 | - | 1 | - | - | - | - |
Розовые Rosaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Сложноцветные Asteracea | 1 | 1 | - | - | 2 | 2 | 1 | 1 |
Фиалковые Violaceae | - | 1 | - | 1 | - | - | - | - |
Хвощевые Equisetaceae | - | - | - | 1 | - | 1 | - | 1 |
Всего видов | 8 | 7 | 6 | 7 | 11 | 6 | 7 | 4 |
Среднеактивные виды | ||||||||
Аспидиевые Aspidiaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Бобовые Fabaceae | - | 1 | - | 1 | 1 | 1 | 1 | 1 |
Вересковые Ericaceaе | - | 1 | - | 1 | 1 | 1 | 1 | 1 |
Ворсянковые Dipsacaceae | 1 | - | 1 | - | - | - | - | - |
Гераниевые Geraniaceae | - | - | - | - | - | 1 | - | 1 |
Грушанковые Pyrolaceae | - | 1 | - | 1 | 1 | 1 | 1 | 1 |
Зверобойные Guttiferae | - | 1 | - | 1 | - | - | - | - |
Злаковые Poaceae | - | - | - | - | - | 1 | - | 1 |
Зонтичные Apiaceae | - | 1 | - | 1 | 1 | - | 1 | - |
Кочедыжниковые Athyriaceae | 1 | 1 | - | 1 | 1 | 1 | - | - |
Крапивные Urticaceae | - | 1 | - | 1 | - | - | - | - |
Крыжовниковые Grossulariaceae | - | - | - | - | - | 1 | - | - |
Лилейные Liliaceae | - | - | - | - | 1 | - | 1 | - |
Норичниковые Scrophulariaceae | - | 1 | - | 1 | - | - | - | - |
Первоцветные Primulaceae | - | - | - | - | - | 1 | - | 1 |
Розовые Rosaceae | 1 | 1 | 1 | 1 | 1 | - | 1 | - |
Сложноцветные Asteracea | - | - | - | - | 1 | 2 | 1 | 1 |
Хвощевые Equisetaceae | - | 2 | - | 1 | - | - | - | - |
Всего видов | 4 | 12 | 3 | 11 | 9 | 10 | 8 | 7 |
Активные виды | ||||||||
Бобовые Fabaceae | 1 | 1 | 1 | 1 | - | - | - | - |
Вересковые Ericaceaе | 1 | - | 1 | - | 1 | 1 | 1 | 1 |
Ворсянковые Dipsacaceae | - | - | - | - | 1 | - | 1 | - |
Гераниевые Geraniaceae | - | 1 | - | 1 | - | - | - | - |
Зверобойные Guttiferae | 1 | - | 1 | - | - | - | - | - |
Злаковые Poaceae | 2 | 1 | 4 | - | - | - | - | - |
Зонтичные Apiaceae | - | 1 | - | 1 | - | - | - | - |
Кипрейные Onagraceae | 1 | 1 | 1 | 1 | - | 1 | - | 1 |
Кисличные Oxalidaceae | - | 1 | - | 1 | - | 1 | - | 1 |
Кочедыжниковые Athyriaceae | - | - | - | - | - | 1 | - | 1 |
Лилейные Liliaceae | - | - | - | - | - | 1 | - | 1 |
Осоковые Cyperaceae | - | - | - | - | - | 1 | - | 1 |
Розовые Rosaceae | - | 1 | - | 1 | 2 | 2 | 1 | 2 |
Сложноцветные Asteracea | - | - | - | - | 1 | - | 1 | - |
Фиалковые Violaceae | 1 | - | 1 | - | - | - | - | - |
Хвощевые Equisetaceae | 1 | - | 1 | - | 2 | 2 | 2 | 2 |
Всего видов | 8 | 7 | 10 | 6 | 7 | 10 | 6 | 10 |
Высокоактивные виды | ||||||||
Бобовые Fabaceae | 1 | - | 1 | - | - | - | - | - |
Гераниевые Geraniaceae | - | - | - | - | 1 | - | 1 | - |
Злаковые Poaceae | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 |
Кипрейные Onagraceae | - | - | - | - | 1 | - | 1 | - |
Розовые Rosaceae | 2 | 2 | 2 | 2 | 1 | 3 | 1 | 3 |
Всего видов | 5 | 4 | 5 | 4 | 4 | 4 | 4 | 4 |
Итого | 36 | 34 | 33 | 32 | 36 | 35 | 27 | 25 |
Согласно фитоценотическому анализу установлено, что основу сообщества лесных культур в фазе смыкания на дренированном участке составляют высокоактивные виды (1 класс активности) (14…15% от общего количества сосудистых видов ценофлоры) – Deschampsia cespitosa L., Calamagrostis epigeios L., Trifolium pratense L., Rubus saxatilis L., Potentilla erecta (L.) Raeusch. Данные виды широко представлены на всей лесокультурной площади и имеют высокий показатель обилия.
С возрастом древостоя в фазе чащи лесных культур, с изменением освещенности и других микроклиматических факторов местообитания, высокоактивные светолюбивые виды напочвенного покрова Trifolium pratense L. и Potentilla erecta (L.) Raeusch. переходят соответственно во 2 и 3 класс активности, а семигелиофитный вид Rubus idaeus L., наоборот, становится высокоактивным видом. Тем самым, к фазе чащи лесных культур общее количество высокоактивных видов сократилось с 5 до 4, при этом на смену Trifolium pratense L. и Potentilla erecta (L.) Raeusch. пришел Rubus idaeus L.
В условиях временного избыточного увлажнения сохраняет свое доминирующее положение Calamagrostis epigeios L., Rubus saxatilis L., а также с учетом обозначенных почвенных особенностей высокую позицию в фазе смыкания занимает Geranium sylvaticum L. и Chamerion angustifolium (L.) Holub., таким образом, эти виды соответствуют 1 классу активности (11…15% от общего количества сосудистых видов ценофлоры).
В фазе чащи древостоя высокоактивные лугово-лесные виды Geranium sylvaticum L. и Chamerion angustifolium (L.) Holub. переходят соответственно во 2 и 3 класс активности, а лесные виды Filipendula ulmaria (L.) Maxim, Rosa acicularis Lindl., наоборот, увеличивают ландшафтное распространение и становятся высокоактивными.
Группу активных видов (2 класс активности) наряду с высокоактивными, мы также относим к наиболее приспособленным к условиям фитоценозов несмотря на то, что они представлены не на всех учетных площадках.
По нашим данным, их доля в популяции составляет 19…40%. Большинство высокоактивных и активных видов относятся к лесной и луговой ценотической группе. Если рассматривать обе группы, то растения 1 и 2 класса активности достаточно многочисленны. На их долю приходится 31…56% популяции, что составляет 10…15 видов растений. Можно отметить, что большинство из рассматриваемых видов сохранились на лесокультурной площади с вейниковой вырубки 1999 г.
Среднеактивные виды (3 класс активности) мы относим к промежуточной группе. Данные виды отличаются более узким диапазоном толерантности к экологическим условиям, так, например, Urtica dioica L. и Dryopteris filix-mas (L.) Scohott. являются более требовательными к почвенному питанию. Наши исследования показали, что по прошествии 14 лет на лесокультурной площади сохраняется от 9 до 30% среднеактивных видов сосудистых растений, а к 22-летнему возрасту их доля возрастает до 29…35%.
Обратный характер сукцессии мы наблюдаем в группе малоактивных и неактивных видов (4…5 классы активности), представители которых встречаются в небольшом количестве на отдельных участках, либо единичными растениями. Малоактивные и неактивные виды не имеют серьезного лесохозяйственного значения для лесных культур, но для экосистемы в целом увеличение пула видов повышает продуктивность системы, улучшает почвенные показатели. В фазе смыкания на данные группы растений приходится значительная часть спектра – 33…53% от общего количества видов ценофлоры. В фазе чащи доля малоактивных и неактивных видов значительно меньше (16…34%) за счет снижения общего видового биоразнообразия из-за высокого распространения активной части флоры.
Таким образом, чем выше участие активных видов в растительном сообществе, тем меньше ресурсов остается малоактивным видам, тем ниже их адаптационный потенциал и общее число видов. Рост участия активных и высокоактивных видов ведет к случайному вытеснению сопутствующих растений. Кроме того, активные виды изменяют условия произрастания для других видов и могут ограничивать их присутствие, исключая из состава ценоза сначала менее устойчивые виды, а затем все более адаптированные до полного их исчезновения.
К малоактивной группе растений напочвенного покрова лесных культур в фазе смыкания отнесены такие временные луговые виды, как Phleum pratense L., Stellaria graminea L., сорно-рудеральные Cirsium arvense (L.) Scop., Urtica dioica L., а также лесные виды Rosa acicularis Lindl., Geranium sylvaticum L., Galium boreale L., Oxalis acetosella L., пока неактивные, но которые спустя десятилетие (в фазе чащи) в большинстве своем займут 2 класс активности.
Со временем в ценофлоре лесных культур фазы чащи исчезло большинство неактивных видов (Achillea millefolium L., Leucanthemum vulgare Lam., Taraxacum officinale Wigg., Gagea lutea (L.) Ker Gawl., Aegopodium podagraria L., Veronica officinalis L.), а им на смену пришли новые виды (Maianthemum bifolium (L.) F.W. Schmidt, Paris quadrifolia L. и др.), не встреченные ранее.
В культурах ели в фазе чащи по причине недостатка освещенности наблюдается существенное сокращение малоактивных и неактивных видов растений живого напочвенного покрова.
4. Заключение
Учитывая вышесказанное, следует подчеркнуть, что происходит постепенное вытеснение луговых видов активной лесной группой растений. В большей степени изменения происходят за счет сокращения менее устойчивых неактивных и малоактивных видов, таким образом, происходит трансформации эколого-ценотической структуры флоры после антропогенного нарушения в направлении восстановления исходной структуры лесной среды.