ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ И ПОЛИМОРФИЗМА ГЕНА DGAT1 НА ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ МОЛОЧНОЙ ПРОДУКТИВНОСТИ И ПРОФИЛЬ ЖИРНЫХ КИСЛОТ МОЛОКА ГОЛШТИНИЗИРОВАННЫХ ЧЕРНО-ПЕСТРЫХ КОРОВ
ВЛИЯНИЕ ФАКТОРОВ СРЕДЫ И ПОЛИМОРФИЗМА ГЕНА DGAT1 НА ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ МОЛОЧНОЙ ПРОДУКТИВНОСТИ И ПРОФИЛЬ ЖИРНЫХ КИСЛОТ МОЛОКА ГОЛШТИНИЗИРОВАННЫХ ЧЕРНО-ПЕСТРЫХ КОРОВ

Аннотация
В статье представлены результаты оценки влияния средовых и генетических факторов, в т.ч. аллельного полиморфизма гена DGAT1 (диацилглицерин-O-ацилтрансфераза) на показатели продуктивности и профиль жирных кислот молочного жира у голштинизированных черно-пестрых коров. Всего было проанализировано поголовье коров объемом 310 голов из опытного хозяйства. Информация о составе молока и продуктивности была представлена записями о ежемесячных контрольных дойках, проводившихся на протяжении двух лет (общее количеством записей n>10000). Отклонений по распределению частот генотипов от закона Харди-Вайнберга не выявлено. У SNP rs109234250 и rs109326954, локализованных в гене DGAT1, преобладал аллель К (0,6). В жирнокислотном составе молочного жира преобладади насыщенные среднецепочечные жирные кислоты. Наследуемость и генетические корреляции исследуемых признаков соответствуют путям образования жирных кислот в организме животного. Наибольшее оценочное значение фиксированного влияния продолжительности лактации (DIM) было выявлено для признаков «содержание жира» и «содержание насыщенных среднецепочечных жирных кислот». Оценка фактора сезона контрольной дойки продемонстрировала максимальное положительное влияние в зимний период с увеличением оценки каждой последующей лактации. Для длинноцепочечных и мононенасыщенных жирных кислот максимальные оценки влияния сезона показаны для весеннего периода и рост до третьей лактации. Полиненасыщенные и транс-изомеры жирных кислот показали отличительные результаты, получившие наименьшие коэффициенты регрессии и максимальные оценки влияния в первую лактацию со снижением в каждой последующей. Максимальное влияние на жирнокислотный состав молочного жира было выявлено для аллеля А гена DGAT1 с преобладанием генотипа АА.
1. Введение
По причине особого влияния на потребительские качества получаемой жиромаслянной продукции профилю жирных кислот молочного жира придается пристальное внимание. За последние десятилетия мнения о вреде или пользе молочного жира менялись на диаметрально противоположные. Это связано с тем, что молочный жир представлен в основном насыщенными жирными кислотами (до 65%), что зачастую ассоциируют с риском возникновения сердечно-сосудистых заболеваний, но в то же время, ряд исследователей сообщает о полезных биологических свойствах олеиновой и стеариновой жирных кислот, что способствуют регуляции уровня липидов в крови. И в дополнение молочный жир содержит такие жирные кислоты как линолевая, альфа-линолевая и коньюгированная линолевая (CLA), полезные с точки зрения питания , , , . Кроме непосредственного влияния молочного жира на здоровье человека при его употреблении, интерес к профилю жирных кислот вызывает его влияние на технологические свойства продуктов переработки молочного жира. Так, увеличение ненасыщенных длинноцепочечных жирных кислот, в противовес короткоцепочечным, увеличивает мягкость и текучесть сливочного масла при низких температурах, но может отрицательно сказаться на вкусовых качествах, по причине усиленного окисления продукта, что также влияет на максимальные сроки хранения .
Состав и содержание жирных кислот молока зависит от путей их синтеза. Условно можно выделить несколько путей образования: непосредственно из рациона, синтез в молочной железе, биогидрирование (бактериальная деградация в рубце) и высвобождение из жировых запасов , . При этом каждому типу жирной кислоты соответствует свой путь синтеза, так насыщенные коротко- и среднецепочечные синтезируются преимущественно de novo в молочной железе, тогда как ненасыщенные и длинноцепочечные жирные кислоты поступают из корма . Регуляция указанных процессов может быть связана со множеством факторов влияющих на них, таких как кормление, порода, номер и стадия лактации. Кормление животного отражается на профиле жирных кислот посредством влияния на регуляцию экспрессии липогенных генов и снижения синтеза de novo и увеличению содержания длинноцепочечных жирных кислот, при повышении содержания полиненасыщенных жирных кислот в кормах. Данные колебания в профиле молочного жира проявляются при смене сезона, т.к. являются отражением смены рациона животных, когда зимой преобладают концентрированные корма и силос, а летом свежая трава, богатая ненасыщенными жирными кислотами . С учетом того, что основная доля жирных кислот синтезируется самим животным, то также стоит брать в расчет и такой фактор как генотип животного, а в частности, полиморфизмы генов регулирующие липидный обмен. Фермент диацилглицерин-O-ацилтрансфераза, кодируемый геном DGAT1 является одним из основных маркеров жирномолочности, т.к. катализирует заключительную стадию биосинтеза триглицеридов. Ассоциации данного гена с содержанием молочного жира и жирных кислот показаны рядом авторов в GWAS – исследованиях, как у молочных пород, так и у мясных , , . Таким образом, целью нашего исследования являлась оценка влияния средовых и генетических факторов на изменчивость продуктивности, а также содержание и состав профиля жирных кислот молочного жира у голштинизированных черно-пестрых коров.
2. Материалы и методы исследования
Образцы ушных выщипов (n=310) для генотипирования и пробы молока были получены от животных голштинизированной черно-пестрой породы (опытное хозяйство ФГБНУ ФИЦ ВИЖ им.Л.К.Эрнста, Краснодарский край). Геномная ДНК выделялась с использованием набора «ДНК-Экстран-2» производства НПО «СИНТОЛ» из выщипов ушных раковин согласно инструкции производителя.
Реакцию амплификации проводили методикой ПЦР в «реальном времени», разработанной Ковальчук С.Н. 2017 г. на приборе Bio-Rad CFX96 в оптимизированных условиях .
В исследовании учитывались следующие показатели: суточный удой (кг), массовая доля жира/белка в молоке (%), содержание казеинов (%), жирнокислотный состав молока (г/100г): С14, С16, С18, С18.1, LCFA, MCFA, MUFA, PUFA, SFA, SCFA, TFA. Данные о фракционном составе молока получали посредством индивидуального отбора проб молока во время проведения ежемесячных контрольных доений – 3 раза в сутки (утро-день-вечер) и последующего анализа в лаборатории селекционного контроля качества молока ФГБНУ ФИЦ ВИЖ им. Л.К. Эрнста на автоматическом анализаторе CombiFoss 7 DC («Foss», Дания) в период с июня 2020 по июль 2022 гг. В исследовательскую базу вошли 10243 наблюдения за период проведения исследования.
Описательные статистические параметры (среднее арифметическое, ошибка, стандартное отклонение) вычислялись при помощи пакета «Анализ данных» в среде MS Excel 2013. Степень изменчивости признаков оценивали по коэффициенту вариации (Cv). Для расчета наследуемости (h2) использовали отношение генетической вариансы к сумме генетической и остаточной варианс. Выявленные различия считали статистически значимыми при p≤0,05.
Оценка влияния генетических маркеров и средовых факторов на продуктивные качества и жирнокислотный состав молока производилась при помощи математической модели смешанного типа, составленной в рамках методологии BLUP Animal Model. Расчёты осуществлялись в программах семейства BLUPF90 . Модель оценки имела следующий вид:
где yijklm – оцениваемый показатель продуктивности или жирнокислотного состава молока; µ – популяционная константа; Li – фиксированный эффект i-й лактации; β – коэффициент линейной регрессии; DIMj – количество дойных дней; Seasonk – сезон лактации, в который был произведен отбор образца молока (Зима-весна-лето-осень); Gl – фиксированный эффект каждого генотипа; animalm – рандомизированный эффект m-ой коровы (n=10243 наблюдения) имеющий нормальное распределение со средней, равной 0, и вариансой, равной σa2 (Vara ~ 0, σa2); eijklm – эффект остатков модели.
3. Основные результаты
Для каждого животного были идентифицированы все генотипы исследуемого гена DGAT1 (таблица 1).
Таблица 1 - Частоты аллелей и генотипов генов DGAT1 у голштинизированного черно-пестрого скота опытного хозяйства
Ген | Частоты генотипов | Частоты аллелей | χ2 | p-value | |||
DGAT1 | AA | AK | KK | A | K | 0,03 | 0,98 |
0,16 | 0,47 | 0,36 | 0,4 | 0,6 | |||
В рамках исследуемой выборки голштинизированных черно-пестрых коров определены частоты встречаемости аллелей А (0,4) и К (0.6) гена DGAT1 SNP rs109234250, rs109326954 с частотой гетерозигот (Ho) 0,47. Отклонений фактического распределения частот генотипов от теоретического согласно закону Харди-Вайнберга не наблюдалось.
Преобладание аллеля К гена DGAT1 SNP rs109234250, rs109326954 в популяциях крупного рогатого скота больше характерно для местных и аборигенных пород скота. Так, у аборигенного африканского крупного рогатого скота частота встречаемости аллеля К составляет от 0,77 до 0,929, в то время как у их помесей с голштинской породой от 0,386 до 0,658 , . Аналогичные данные показаны для черного японского мясного скота, где минорным являлся аллель А с низкой частотой встречаемости генотипа АА – 0,056 . Среди отечественных пород скота повышение частоты встречаемости аллеля К принято ассоциировать с прилитием голштинской крови, особенно у черно-пестрой голштинизированной породы , а также с высокой частотой гетерозигот в различных популяциях холмогорской породы от 0,35 до 0,49 , . Для европейской популяции голштинской породы характерно незначительное преобладание аллеля А, с долей гетерозигот – 0,47 , , , , приближенная ситуация показана и для китайской популяции голштинского скота, с тем исключением, что доля гомозиготного генотипа АА и доля гетерозигот были одинаковы – 0,479 и минимальным присутствием генотипа КК – 0,042 .
Из литературных данных также показано преобладание аллеля А гена DGAT1 в иностранных популяциях мясных и молочных породах крупного рогатого скота, при этом сопоставимые данные получены для таких европейских пород скота как симментальская, бурая, шароле, лимузин, абердин-ангуская, герефорд, у которых частота встречаемости аллеля А превышала 0,8, а иногда наблюдалось полное отсутствие аллеля K в популяции , , , .
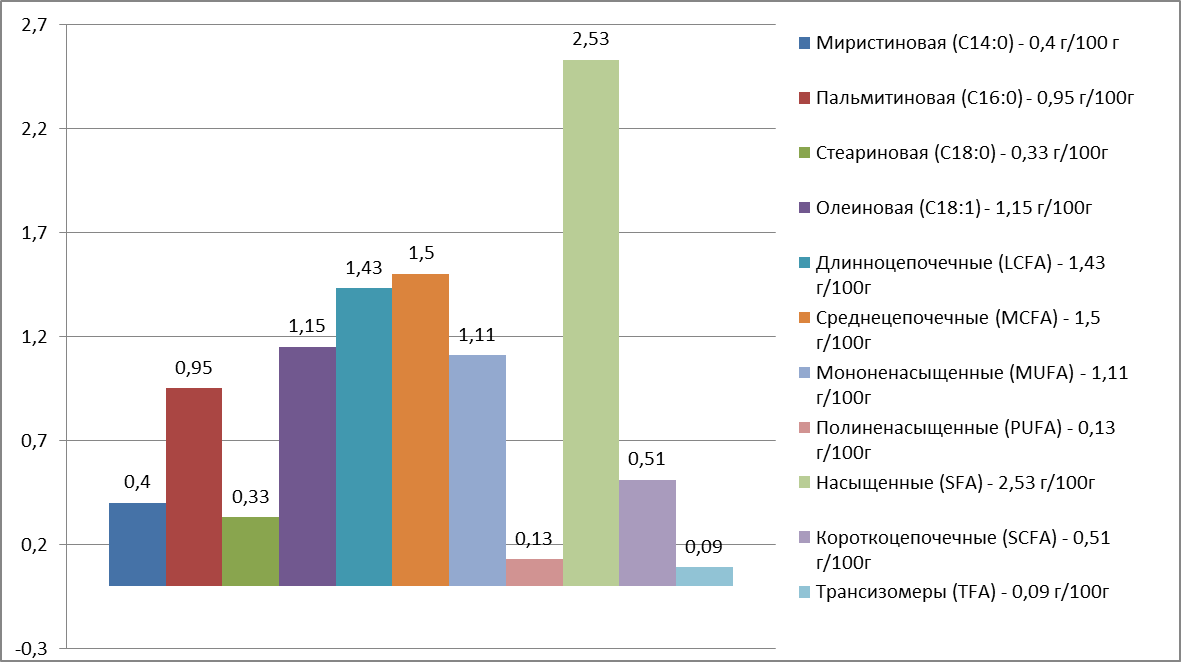
Содержание жирных кислот в молоке голштинизированных черно-пестрых коров опытного хозяйства
Наследуемость показателей продуктивности и жирнокислотного состава молока колебалась в диапазоне от 0,13 до 0,58 (табл. 2). Для длинноцепочечных (LCFA), мононенасыщенных (MUFA), стеариновой (С18:0) и олеиновой (С18:1) жирных кислот были получены наименьшие коэффициенты наследуемости – 0,13-0,19. Максимальные коэффициенты были получены для миристиновой (С14:0) – 0,58, пальмитиновой (С16:0) – 0,41, среднецепочечных (MCFA) – 0,47. Полученные нами результаты согласуются с рядом авторов, также описавших высокую степень наследуемости – свыше 0,4 для С14:0 и С16:0 , , что может быть связано с тем, что молочный жир до 65% представлен насыщенными жирными кислотами, в частности, миристиновой и пальмитиновой, а также путями синтеза жирных кислот. Данные результаты подтверждаются очень высокой генетической корреляцией между жирными кислотами и массовой долей жира: МДЖ%/С14:0 – 0,87, МДЖ%/С16:0 – 0,94, МДЖ%/MCFA – 0,94, МДЖ%/SFA – 0,97. Также очень высокая положительная корреляция была получена для длинноцепочечных жирных кислот (LCFA) и стеариновой (0,89), олеиновой (0,97) жирных кислот; мононенасыщенных жирных кислот и стеариновой (0,78), олеиновой (0,96), длинноцепочечных жирных кислот (0,93). Для полиненасыщенных и транс-изомеров жирных кислот, была также получена очень высокая положительная корреляция (0,77), т.к. TFA являются побочным продуктом образования насыщенных жирных кислот путем гидрогенизации жирных кислот в рубце животного. Образование транс-изомеров жирных кислот из полиненасыщенных и то, что PUFA представлены в основном длинноцепочечными жирными кислотами подтверждается высокой положительной корреляцией между LCFA (0,67), С18:0 (0,61), С18:1 (0,69) и PUFA, а также средней положительной с TFA (0,38/0,46/0,34), низкой корреляцией между TFA/MUFA – 0,26, отрицательной корреляцией между PUFA и C14:0 (-0,19), C16:0 (-0.02), MCFA (-0,06) и средней отрицательной между TFA/C14:0 (-0,41), TFA/C16:0 (-0.33), TFA/MCFA (-0,37). Также низкая отрицательная корреляция между TFA и содержанием жира в молоке и остальными группами жирных кислот может подтверждать влияние трансизомеров ЖК на экспрессию генов, ассоциированных с синтезом молочного жира .
Таблица 2 - Наследуемость h2 (по диагонали) и генетическая корреляция (под диагональю) между показателями продуктивности и жирнокислотным составом молока
| Сут. удой | МДЖ% | МДБ% | Cas B | C14:0 | C16:0 | C18:0 | C18:1 | LCFA | MCFA | MUFA | PUFA | SFA | SCFA | TFA |
Суточный удой | 0,39 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
МДЖ% | -0,42 | 0,31 |
|
|
|
|
|
|
|
|
|
|
|
|
|
МДБ% | -0,47 | 0,63 | 0,41 |
|
|
|
|
|
|
|
|
|
|
|
|
Cas B | -0,48 | 0,68 | 0,99 | 0,42 |
|
|
|
|
|
|
|
|
|
|
|
C14:0 | -0,46 | 0,87 | 0,61 | 0,68 | 0,58 |
|
|
|
|
|
|
|
|
|
|
C16:0 | -0,42 | 0,94 | 0,56 | 0,60 | 0,89 | 0,41 |
|
|
|
|
|
|
|
|
|
C18:0 | -0,05 | 0,53 | 0,18 | 0,16 | 0,11 | 0,41 | 0,19 |
|
|
|
|
|
|
|
|
C18:1 | -0,21 | 0,66 | 0,43 | 0,41 | 0,25 | 0,48 | 0,83 | 0,13 |
|
|
|
|
|
|
|
LCFA | -0,15 | 0,63 | 0,32 | 0,32 | 0,21 | 0,44 | 0,89 | 0,97 | 0,13 |
|
|
|
|
|
|
MCFA | -0,45 | 0,94 | 0,64 | 0,69 | 0,95 | 0,98 | 0,32 | 0,43 | 0,38 | 0,47 |
|
|
|
|
|
MUFA | -0,30 | 0,77 | 0,47 | 0,48 | 0,43 | 0,59 | 0,78 | 0,96 | 0,93 | 0,56 | 0,13 |
|
|
|
|
PUFA | 0,02 | 0,18 | 0,29 | 0,25 | -0,19 | -0,02 | 0,61 | 0,69 | 0,67 | -0,06 | 0,57 | 0,22 |
|
|
|
SFA | -0,37 | 0,97 | 0,62 | 0,66 | 0,82 | 0,96 | 0,56 | 0,64 | 0,61 | 0,95 | 0,71 | 0,20 | 0,33 |
|
|
SCFA | -0,27 | 0,91 | 0,62 | 0,65 | 0,72 | 0,85 | 0,58 | 0,66 | 0,64 | 0,84 | 0,68 | 0,37 | 0,95 | 0,30 |
|
TFA | 0,15 | -0,14 | -0,05 | -0,09 | -0,41 | -0,33 | 0,46 | 0,34 | 0,38 | -0,37 | 0,26 | 0,77 | -0,16 | -0,03 | 0,24 |
Влияние продолжительности лактации, выраженное через коэффициент регрессии измеряемого признака на количество дойных дней, представлено в таблице 3. Количество дойных дней варьировало в диапазоне от 10 до 1184. Полученные результаты соответствуют изменениям в продуктивности животного в течение лактации, так коэффициент регрессии для удоя составил -0,028, что говорит о естественном уменьшении продуктивности к концу лактации. Напротив, максимальные значения были получены для массовой доли жира и белка и, соответственно, содержания казеинов, а также для среднецепочечных и насыщенных жирных кислот, как максимально представленных в молочном жире.
Таблица 3 - Коэффициент регрессии показателя количество дойных дней (DIM)
Показатель | Коэффициент регресии |
Суточный удой | -0,028±0,0004 |
МДЖ% | 0,002±0,00007 |
МДБ% | 0,002±0,00002 |
Казеин | 0,002±0,00002 |
Миристиновая (С14:0) | 0,0003±0,000006 |
Пальмитиновая (С16:0) | 0,0005±0,00002 |
Стеариновая (С18:0) | -0,00002±0,00001 |
Олеиновая (С18:1) | 0,0003±0,00002 |
Длинноцепочечные (LCFA) | 0,0002±0,00003 |
Среднецепочечные (MCFA) | 0,001±0,00003 |
Мононенасыщенные (MUFA) | 0,0003±0,00002 |
Полиненасыщенные (PUFA) | 0,00003±0,000002 |
Насыщенные (SFA) | 0,001±0,00004 |
Короткоцепочечные (SCFA) | 0,0002±0,00001 |
Трансизомеры (TFA) | -0,00002±0,000003 |
Стоит отметить, что в ранний период лактации у молочных коров часто наблюдается отрицательный энергетический баланс, что компенсируется активацией резервов организма, а именно запасов собственной жировой ткани, что отражается в профиле жирных кислот и сниженным содержанием коротко- и среднецепочечных жирных кислот , .
Генетические оценки состава молочного жира в зависимости от сезона лактации приведены в таблице 4. Было установлено наибольшее отрицательное влияние летнего периода на продуктивные качества и состав молочного жира коров. Так же отрицательные показатели были получены для насыщенных (SFA), среднецепочечных (MCFA), короткоцепочечных (SCFA) и пальмитиновой (С16:0) с миристиновой (С14:0) жирных кислот в весенний период. Для тех же групп и индивидуальных жирных кислот, а также массовой доли жира и белка было показано максимальное положительное влияние в зимний период. Также зимний период лактации положительно влиял на содержание стеариновой (С18:0), длиноцепочечных (LCFA) и ненасыщенных жирных кислот, но наибольшие значения были показаны для весеннего периода. Различий во влиянии зимнего и весеннего периодов на содержание олеиновой (С18:1) жирной кислоты обнаружено не было. Для транс-изомеров (TFA) жирных кислот положительное влияние было выявлено только в весенний период лактации, также как и для суточного удоя. Данные о снижении содержания насыщенных коротко- и среднецепочечных жирных кислот в летний период и рост в зимний полностью согласуются с литературными данными, но отличные для насыщенных и ненасыщенных длинноцепочечных жирных кислот, где был показан их рост именно в летний период. Данная закономерность была объяснена авторами, как влияние наличия свежескошенной травы в рационе питания, в которой преобладают ненасыщенные жирные кислоты или выпас животных .
Таблица 4 - Влияние сезона лактации на показатели продуктивности и жирнокислотный состав молока
Показатель | Весна | Зима | Лето | Осень |
Суточный удой | 0,666±0,138 | -1,162±0,152 | -1,218±0,15 | 0,0 |
МДЖ% | -0,099±0,024 | 0,18±0,026 | -0,679±0,026 | 0,0 |
МДБ% | -0,088±0,008 | -0,013±0,009 | -0,218±0,009 | 0,0 |
Казеин | -0,084±0,007 | -0,01±0,008 | -0,223±0,008 | 0,0 |
Миристиновая (С14:0) | -0,031±0,002 | 0,017±0,002 | -0,068±0,002 | 0,0 |
Пальмитиновая (С16:0) | -0,048±0,006 | 0,059±0,006 | -0,139±0,006 | 0,0 |
Стеариновая (С18:0) | 0,015±0,003 | 0,009±0,003 | -0,046±0,003 | 0,0 |
Олеиновая (С18:1) | 0,03±0,008 | 0,03±0,009 | -0,134±0,009 | 0,0 |
Длинноцепочечные (LCFA) | 0,046±0,011 | 0,036±0,012 | -0,187±0,012 | 0,0 |
Среднецепочечные (MCFA) | -0,096±0,009 | 0,087±0,009 | -0,260±0,009 | 0,0 |
Мононенасыщенные (MUFA) | 0,032±0,008 | 0,029±0,009 | -0,106±0,009 | 0,0 |
Полиненасыщенные (PUFA) | 0,007±0,001 | 0,001±0,001 | -0,014±0,001 | 0,0 |
Насыщенные (SFA) | -0,122±0,015 | 0,152±0,017 | -0,519±0,017 | 0,0 |
Короткоцепочечные (SCFA) | -0,027±0,003 | 0,032±0,004 | -0,131±0,004 | 0,0 |
Трансизомеры (TFA) | 0,017±0,001 | -0,001±0,001 | -0,002±0,001 | 0,0 |
Результаты оценки влияния номера лактации на показатели продуктивности и состав жирных кислот демонстрируют общую закономерность в изменении обильномолочности и жирномолочности в зависимости от номера лактации (таблица 5).
Таблица 5 - Влияние номера лактации на показатели продуктивности и жирнокислотный состав молока
Номер лактации | 1 | 2 | 3 | 4 | 5 |
Суточный удой | 2,632±0,578 | 2,983±0,57 | 1,880±0,58 | 0,983±0,644 | 0,0 |
МДЖ% | -0,69±0,086 | -0,6±0,084 | -0,45±0,087 | -0,092±0,099 | 0,0 |
МДБ% | -0,13±0,035 | -0,10±0,034 | -0,03±0,035 | -0,034±0,038 | 0,0 |
Казеин | -0,12±0,032 | -0,09±0,031 | -0,02±0,032 | -0,017±0,035 | 0,0 |
Миристиновая (С14:0) | -0,15±0,012 | -0,11±0,012 | -0,07±0,012 | -0,018±0,013 | 0,0 |
Пальмитиновая (С16:0) | -0,19±0,025 | -0,14±0,024 | -0,12±0,025 | 0,005±0,027 | 0,0 |
Стеариновая (С18:0) | 0,004±0,008 | -0,004±0,01 | -0,02±0,008 | -0,003±0,010 | 0,0 |
Олеиновая (С18:1) | -0,008±0,020 | -0,04±0,020 | -0,07±0,021 | -0,03±0,026 | 0,0 |
Длинноцепочечные (LCFA) | 0,04±0,027 | -0,02±0,027 | -0,07±0,028 | -0,03±0,035 | 0,0 |
Среднецепочечные (MCFA) | -0,4±0,041 | -0,28±0,040 | -0,21±0,041 | -0,023±0,045 | 0,0 |
Мононенасыщенные (MUFA) | -0,06±0,019 | -0,09±0,019 | -0,09±0,020 | -0,035±0,025 | 0,0 |
Полиненасыщенные (PUFA) | 0,017±0,002 | 0,012±0,002 | 0,01±0,002 | 0,002±0,003 | 0,0 |
Насыщенные (SFA) | -0,31±0,058 | -0,22±0,057 | -0,19±0,059 | 0,021±0,066 | 0,0 |
Короткоцепочечные (SCFA) | -0,03±0,012 | -0,02±0,012 | -0,02±0,012 | 0,011±0,014 | 0,0 |
Трансизомеры (TFA) | 0,013±0,003 | 0,008±0,003 | 0,004±0,003 | -0,001±0,004 | 0,0 |
Показано, что оценка массовой доли жира, а также насыщенных среднецепочечных жирных кислот, увеличивается с каждой последующей лактацией, достигая максимума в 4-ую (пальмитиновая, насыщенные жирные кислоты) или 5-ую лактацию (миристиновая, среднецепочечные жирные кислоты). Влияние на количество длинноцепочечных жирных кислот, существенно отличалось от среднецепочечных, так, максимальная оценка влияния номера лактации продемонстрирована для 1-ой лактации и к 3-ей достигает минимума. Максимально приближенные результаты были показаны и у мононенасыщенных жирных кислот, с той разницей, что максимально отрицательные оценки были получены для 2-ой и 3-ей лактации. Особый интерес представляют полиненасыщенные и транс-изомеры жирных кислот, т.к. для них показаны практически одинаковые тенденции и изменения оценок в зависимости от лактации.
Данные показатели и взаимодействия полностью согласуются с выше полученными данными, особенно в контексте объяснения путей получения жирных кислот молочного жира. Так, для первотелок характерен жирнокислотный профиль с преобладанием ненасыщеных жирных кислот. Авторы это связывают с тем, что первотелки все еще развиваются по сравнению со взрослыми коровами, также была отмечена сниженная экспрессия комплекса ферментов синтетаз жирных кислот в молочной железе первотелок, которая выходит на уровень, характерный для коров последующих лактаций, только к последней трети первой лактации. Таким образом, происходит активное использование жирных кислот из кормов и собственных резервов , , .
Влияние аллельного полиморфизма гена DGAT1 на молочную продуктивность и жирнокислотный состав молока представлено в таблице 6.
Таблица 6 - Оценки влияния генотипов SNP rs109234250, rs109326954 гена DGAT1 на показатели молочной продуктивности и профиль жирных кислот
Показатель | DGAT1 | ||
AA | AK | KK | |
Суточный удой | -1,674±1,089 | -0,658±0,795 | 0,0 |
МДЖ% | 0,54±0,136 | 0,326±0,099 | 0,0 |
МДБ% | 0,234±0,064 | 0,12±0,047 | 0,0 |
Казеин | 0,215±0,054 | 0,117±0,042 | 0,0 |
Миристиновая (С14:0) | 0,052±0,017 | 0,036±0,012 | 0,0 |
Пальмитиновая (С16:0) | 0,151±0,038 | 0,092±0,028 | 0,0 |
Стеариновая (С18:0) | 0,033±0,012 | 0,017±0,009 | 0,0 |
Олеиновая (С18:1) | 0,125±0,03 | 0,064±0,022 | 0,0 |
Длинноцепочечные (LCFA) | 0,148±0,041 | 0,083±0,03 | 0,0 |
Среднецепочечные (MCFA) | 0,237±0,06 | 0,148±0,044 | 0,0 |
Мононенасыщенные (MUFA) | 0,107±0,029 | 0,058±0,021 | 0,0 |
Полиненасыщенные (PUFA) | 0,011±0,003 | 0,006±0,002 | 0,0 |
Насыщенные (SFA) | 0,406±0,092 | 0,243±0,067 | 0,0 |
Короткоцепочечные (SCFA) | 0,09±0,02 | 0,054±0,014 | 0,0 |
Транс-изомеры (TFA) | 0,004±0,005 | 0,003±0,003 | 0,0 |
Влияние полиморфизма rs109234250, rs109326954 гена DGAT1 на молочную продуктивность и состав жирных кислот показало положительную ассоциацию для всех рассматриваемых показателей для аллеля А, кроме удоя, где желательным аллелем является аллель К. Полученные результаты отличались от имеющихся данных, так для разводимых в нашей стране пород скота в основном было показано положительное влияние аллеля А на обильномолочность, в то время как на жирномолочность больше влиял аллель К , , , но обратный результат был получен для голштинизированных симменталов . Среди иностранных исследователей, также имеются противоречивые результаты, вплоть до отсутствия достоверных ассоциаций. Однозначного влияние какого-либо из аллеля гена DGAT1 на жирнокислотный состав молочного жира также не представлено. Так, положительное влияние аллеля А, показанное E.Samková et al. 2021 для насыщенных коротко- и среднецепочечных жирных кислот и положительное влияние аллеля К на ненасыщенные жирные кислоты, не согласуются с ранее полученными данными A.M. Carvajal et al 2015 и Schennink et al. 2008 , Ассоциация аллеля К с длинноцепочечными жирными кислотами, у последних авторов, отличалась от результатов G. Conte et al. 2010 , который показал взаимосвязь с аллелем А.
Влияние аллеля А на содержание транс-изомеров жирных кислот хоть и было показано, но достоверных отличий обнаружено не было.
4. Заключение
В нашем исследовании было изучено влияние на молочную продуктивность и состав жирных кислот таких факторов, как возраст животного, выраженный в номере лактации, продолжительность и сезон лактации, а также полиморфизм гена DGAT1. Полученные результаты предполагают возможность изменения профиля жирных кислот молочного жира в результате отбора и селекции. Но стоит учитывать, что селекционная работа в равной степени может регулироваться скорректированным рационом кормления с содержанием желательных жирных кислот. Использование информации о генотипах гена DGAT1 в полной мере на данный момент затруднительно, т.к. наибольшее влияние аллельного полиморфизма показано для содержания жира в молоке, что, возможно, корректирует оценки жирнокислотного состава, но однозначных сведений о влиянии определенного аллеля нет. Это свидетельствует о необходимости дополнительных дальнейших исследований в данном направлении с расширением исследовательской базы и параметров.