Генетическая вариабельность эвенской породы домашних северных оленей Усть-Янской популяции
Генетическая вариабельность эвенской породы домашних северных оленей Усть-Янской популяции
Аннотация
Статья посвящена исследованию генетической структуры эвенской породы домашних северных оленей Усть-Янской популяции Республики Саха (Якутия). Оленеводство является важной отраслью экономики региона, обеспечивающей продовольственную безопасность и социальную стабильность населения. Несмотря на сокращение общего поголовья оленей в России и Якутии, в Усть-Янском районе отмечен значительный прирост численности этих животных. Исследование генетического разнообразия имеет ключевое значение для поддержания продуктивности и адаптивности породы к суровым климатическим условиям. В работе использовались современные методы молекулярной генетики, включая мультиплексный ПЦР-анализ по 16 микросателлитным локусам. Результаты показали высокий уровень генетического разнообразия в исследуемой популяции, наличие уникальных приватных аллелей и сходство с другими популяциями региона. Полученные данные могут способствовать разработке эффективных программ селекции и сохранению биоразнообразия северных домашних оленей.
1. Введение
Северное домашнее оленеводство продолжает оставаться ключевым направлением экономической активности и образом жизни коренных малочисленных народов Севера, Сибири и Дальнего Востока, выполняя важные социальные функции, такие как обеспечение продовольственной безопасности, поддержка социальной стабильности в сельских районах, борьба с бедностью и помощь коренному населению в преодолении сложных экономических условий. Однако развитие этой отрасли в Республике Саха (Якутия) сталкивается с рядом трудностей, включая слабую организацию селекционной работы, сокращение численности северных домашних оленей, ухудшение показателей выхода телят и сохранения взрослых особей , , .
По сравнению с 1990 годом поголовье северных домашних оленей в России сократилось на 29,6% (с 2260,6 до 1592,4 тыс. голов), в Якутии – сократилось на 52,5% (с 361,6 до 171,6 тыс. голов). В Якутии численность домашних оленей неуклонно сокращалось с 2011 по 2015 гг. по данным на конец года. За последние пять лет (2019-2023 гг.) по республике отмечается рост поголовья северных домашних оленей на 12,82% (с 152,1 до 171,6 тыс. голов). Однако, по статистическим данным за 2019-2022 гг., в Усть-Янском районе РС (Я) на начало 2024 г. по сравнению с 2020 г. наблюдается увеличение поголовья северных домашних оленей на 27,6% (с 20583 до 32756 голов).
Оценка генетического разнообразия пород домашних животных имеет важное значение для поддержания их продуктивных характеристик и адаптации к изменяющимся условиям окружающей среды , , , , . В частности, изучение генетической вариабельности у домашних северных оленей эвенской породы представляет особый интерес, так как эта порода традиционно разводится в суровых климатических условиях Севера и играет ключевую роль в экономике и культуре коренных народов региона.
Для оценки генетического разнообразия домашних северных оленей эвенской породы используются различные молекулярно-генетические методы, включая анализ полиморфизма ДНК, секвенирование генома и изучение микросателлитных локусов. Эти методы позволяют выявить уровень гетерозиготности, наличие редких аллелей и степень инбридинга в популяциях. Например, в работе Н.П. Филипповой и соавторов (2020) было проведено исследование генетического разнообразия домашних северных оленей эвенской породы по локусам трансферрина и микросателлитов . Результаты показали что эвенская порода оленей, разводимая в Булунском районе, имеет свои характерные генетические особенности. Аналогичные результаты были получены в других исследованиях, где использовался метод микросателлитного анализа (А.Д. Соловьева и др., 2022) .
Целью данного исследования является оценка генетического разнообразия и генетической структуры популяций эвенкийской породы северного оленя методом анализа микросателлитов.
2. Методы и принципы исследования
В ходе выполнения работы использованы классические методы и передовые подходы, основанные на методах молекулярной генетики. Для характеристики генетических особенностей был отобран генетический материал от молодняка текущего года (n=78). Лабораторные исследования проведены в лаборатории молекулярно-генетической экспертизы «Племэксперт» ГБУ РС (Я) «Сахаагроплем». Генотипирование проведено набором реагентов для мультиплексного анализа 16 микросателлитных маркеров COrDIS Rangifer, в соответствии с протоколом производителя (ООО «Гордиз», Россия), амплификация материала на термоциклере "Т100" (Bio-Rad, США), микросателлитный профиль получен, используя генетический секвенатор «Нанофор-05» с лазерным детектором (ООО «Синтол», Россия). Обработка данных произведена с использованием надстройки для Microsoft Excel "GeneAlex 6.51" (США).
3. Результаты и обсуждение
Исследование генетической структуры популяций домашних северных оленей является важным аспектом в понимании их эволюции, адаптации к различным экологическим условиям и разработки стратегий управления генетическими ресурсами. Одним из ключевых методов оценки генетического разнообразия является анализ частоты встречаемости аллелей микросателлитных локусов. Частоты встречаемости аллелей микросателлитных локусов эвенской породы домашних северных оленей Усть-Янской популяции представлены в таблице 1. Анализ показал значительную вариацию частот встречаемости аллелей среди различных локусов. Некоторые аллели, такие как 154 в локусе BMS1788, 216 в локусе NVHRT16, 205 в локусе Rt30, 180 в локусе C143, 202 в локусе Rt30, 215 в локусе C217 и 267 в локусе T40, демонстрируют значительную частоту встречаемости. Это может свидетельствовать о доминировании этих аллелей в популяции. Полученные результаты указывают на высокую степень генетического разнообразия в исследуемой популяции.
Таблица 1 - Частота встречаемости аллелей микросателлитных локусов эвенской породы домашних северных оленей Усть-Янской популяции
Локус | Аллель | Частота | Локус | Аллель | Частота | Локус | Аллель | Частота |
BMS1788 | 144 | 0,173 | NVHRT16 | 206 | 0,256 | Rt30 | 205 | 0,340 |
146 | 0,122 | 208 | 0,109 | 207 | 0,103 | |||
152 | 0,058 | 214 | 0,154 | 209 | 0,103 | |||
154 | 0,391 | 216 | 0,359 | 211 | 0,186 | |||
155 | 0,038 | 226 | 0,071 | 219 | 0,064 | |||
156 | 0,071 | OheQ | 284 | 0,083 | 223 | 0,109 | ||
BMS745 | 130 | 0,058 | 295 | 0,058 | Rt6 | 190 | 0,071 | |
132 | 0,359 | 306 | 0,250 | 198 | 0,064 | |||
133 | 0,077 | 307 | 0,103 | 200 | 0,115 | |||
134 | 0,237 | 311 | 0,179 | 202 | 0,333 | |||
136 | 0,218 | 318 | 0,096 | 204 | 0,090 | |||
C143 | 176 | 0,269 | 323 | 0,115 | 206 | 0,263 | ||
180 | 0,679 | 247 | 0,333 | Rt7 | 238 | 0,083 | ||
184 | 0,051 | 249 | 0,090 | 242 | 0,346 | |||
C276 | 354 | 0,205 | 251 | 0,128 | 244 | 0,256 | ||
414 | 0,109 | 253 | 0,141 | 250 | 0,058 | |||
418 | 0,051 | Rt1 | 259 | 0,071 | 252 | 0,115 | ||
430 | 0,359 | 261 | 0,071 | 254 | 0,128 | |||
434 | 0,269 | 263 | 0,071 | Rt9 | 133 | 0,135 | ||
C32 | 298 | 0,141 | Rt24 | 236 | 0,282 | 143 | 0,071 | |
300 | 0,051 | 240 | 0,115 | 145 | 0,154 | |||
306 | 0,423 | 244 | 0,083 | 147 | 0,077 | |||
322 | 0,385 | 248 | 0,103 | 151 | 0,090 | |||
FCB193 | 126 | 0,071 | 252 | 0,179 | 153 | 0,212 | ||
128 | 0,506 | 256 | 0,090 | 155 | 0,186 | |||
130 | 0,058 | 268 | 0,051 | C217 | 215 | 0,872 | ||
134 | 0,058 | T40 | 259 | 0,135 | 219 | 0,128 | ||
136 | 0,141 | 267 | 0,699 |
| ||||
138 | 0,128 | 271 | 0,096 | |||||
Примечание: (n=78)
Основные популяционные генетические характеристики представлены в таблице 2. Среднее количество аллелей на локус (Na) составило 8,563, эффективное число аллелей (Ne) – 4,308, наблюдаемая гетерозиготность (Ho) – 0,699, ожидаемая гетерозиготность (He) – 0,708, а индекс фиксации (F) – 0,013. Эти данные свидетельствуют о высоком уровне генетического разнообразия в исследуемой популяции.
Таблица 2 - Популяционно генетическая характеристика Усть-Янской популяции
Локус | Na | Ne | Ho | He | F |
Среднее | 8,563 | 4,308 | 0,699 | 0,708 | 0,013 |
Примечание: (n=78)
При сравнении показателей генетического разнообразия Усть-Янской популяции с показателями Оймяконской и Томпонской популяций
, было обнаружено некоторое различие между ними. Так, среднее количество аллелей на локус (Na) в Усть-Янской популяции оказалось выше, чем в Томпонской, но ниже, чем в Оймяконской. Однако различия в других показателях оказались незначительными, что говорит о схожем уровне генетического разнообразия всех трех популяций (см. рис. 1).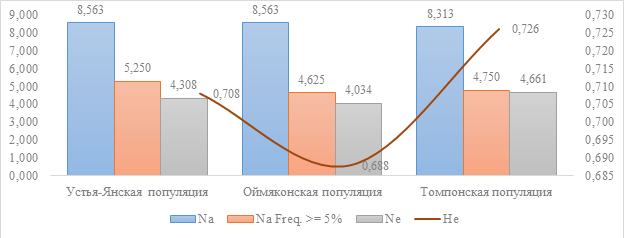
Рисунок 1 - Сравнение с Оймяконской и Томпонской популяцией
Особое внимание следует уделить приватным аллелям, обнаруженным в изученной популяции. Присутствие таких аллелей может свидетельствовать об уникальности генетического состава данной популяции и ее способности к адаптации в специфичных условиях обитания. В нашем исследовании было выявлено 15 приватных аллелей, частота которых варьировалась от 0,006 до 0,051 (табл. 3).
Таблица 3 - Приватные алллели микросателлитных локусов у домашних северных оленей Усть-Янской популяции
№ | Локус | Аллель, п.н. | Частота |
1 | BMS1788 | 141 | 0,013 |
2 | BMS1788 | 170 | 0,013 |
3 | BMS745 | 122 | 0,006 |
4 | BMS745 | 124 | 0,006 |
5 | C276 | 426 | 0,006 |
6 | C32 | 300 | 0,051 |
7 | OheQ | 280 | 0,006 |
8 | OheQ | 299 | 0,006 |
9 | OheQ | 310 | 0,026 |
10 | Rt24 | 268 | 0,051 |
11 | Rt7 | 232 | 0,006 |
12 | Rt7 | 234 | 0,006 |
13 | Rt9 | 135 | 0,013 |
14 | T40 | 326 | 0,006 |
15 | T40 | 333 | 0,006 |
Примечание: (n=78)
Анализ генетического расстояния по Нею показал наименьшее генетическое расстояние (0,051) между Устья-Янской и Томпонской популяциями, что свидетельствует об относительно высоком уровне генетического сходства между этими группами, тогда как тогда как расстояние с Оймяконской популяцией было несколько выше (0,051). В то же время, дистанция между Оймяконской и Томпонской популяциями составила 0,086, что говорит о значительном генетическом отличии этих двух групп (табл. 4). Это может указывать на наличие общих генетических корней у этих популяций, однако также свидетельствует о некотором уровне дифференциации, обусловленном различными условиями среды и историей разведения.
Таблица 4 - Генетическое расстояние между популяциями по Нею
Устья-Янская популяция (n=78) | Оймяконская популяция (n=349) | Томпонская популяция (n=47) | |
0,000 | – | – | Устья-Янская популяция (n=78) |
0,057 | 0,000 | – | Оймяконская популяция (n=349) |
0,051 | 0,086 | 0,000 | Томпонская популяция (n=47) |
Полученные результаты позволяют сделать вывод о том, что домашние северные олени эвенской породы обладают высоким уровнем генетического разнообразия, что обеспечивает им устойчивость к изменениям условий окружающей среды и поддерживает их продуктивность. Важно отметить, что присутствие приватных аллелей подчеркивает уникальность генетического фонда данной популяции и требует особого внимания со стороны специалистов по селекции и сохранению биоразнообразия.
4. Заключение
Оценка генетического разнообразия домашних северных оленей эвенской породы является важной задачей для сохранения и улучшения их продуктивных качеств. Проведенное нами исследование позволило выявить внутрипородную дифференциацию популяций домашних северных оленей эвенской породы и оценить уровень генетического разнообразия породы. Полученные результаты позволят повысить эффективность племенных работ путём улучшения методов племенной оценки и контроля над селекционными процессами. Кроме того, результаты могут быть использованы для разработки селекционных программ, направленных на сохранение и улучшение генетического потенциала эвенской породы домашних северных оленей.