СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ГЕНЕТИЧЕСКИХ ПРОФИЛЕЙ СОВРЕМЕННЫХ ПРЕДСТАВИТЕЛЕЙ ОРЛОВСКОЙ РЫСИСТОЙ ПОРОДЫ И ИХ ПРЕДКОВ
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ГЕНЕТИЧЕСКИХ ПРОФИЛЕЙ СОВРЕМЕННЫХ ПРЕДСТАВИТЕЛЕЙ ОРЛОВСКОЙ РЫСИСТОЙ ПОРОДЫ И ИХ ПРЕДКОВ
Аннотация
Целью данного исследования была сравнительная характеристика генетических профилей современных представителей лошадей орловской рысистой породы, полученной в разных конных заводах, с образцами исторической ДНК, полученной из скелетов лошадей той же породы, существовавших более 100 лет назад.
Для анализа были использованы данные генотипирования по 17 STR-маркерам.
Были получены препараты ДНК из музейных образцов тканей лошадей орловской рысистой породы – Бычка (1824 г.р.) и Улова (1928 г.р.). Произведено STR-генотипирование музейных образцов и образцов современных представителей орловской рысистой породы (73 головы). Оценка популяционно-генетических характеристик современного поголовья орловской рысистой породы лошадей с учетом компонентов предковых форм показала, что достаточно большая доля современных представителей орловской рысистой породы значительно отличается от своих древних представителей.
1. Введение
Орловская рысистая порода лошадей является первой заводской породой лошадей, созданной в России. С момента ее создания прошло почти два столетия. За это время порода прошла путь от чистопородного разведения до скрещивания с американскими рысаками и рядом других пород до признания того факта, что она национальным достоянием и представляет собой культурно-историческую ценность.
С одной стороны, в настоящее время орловская рысистая порода не входит в «группу риска» по классификации ФАО: маточное поголовье увеличивается, к ипподромным испытаниям добавляется спортивно-досуговое направление, что ведет к увеличению численности лошадей этой породы, применяемых также в конном спорте. С другой стороны, каждая порода имеет свои популяционно-генетические характеристики, сформировавшиеся в результате племенной работы с ней и отличающие ее от других пород того же направления продуктивности
, , .Поэтому при идентификации принадлежности лошадей к орловской рысистой породе становится значимым использовать при оценке не только данные STR-генотипирования лошадей, живущих в настоящее время, но и данные животных, имеющих предковые генетические компоненты, характерные для поголовья этой породы сразу после создания, когда основным методом разведения являлось чистопородное разведение
, , , . В настоящее время база данных микросателлитных профилей, на основании которой осуществляется оценка достоверности происхождения пробанда и его отнесения к орловской рысистой породе составлена из данных о генотипах лошадей, которые рождены в последние два десятка лет. Поэтому в нее могут входить лошади, сохраняющие в своей структуре геномные компоненты, специфичные для орловского рысака, так и животные, которые имеют компоненты, привнесенные в геном данной породы в процессе ее развития в последние полтора-два столетия .С этой точки зрения извлечение исторической ДНК из останков двух лошадей, являвшихся яркими представителями породами разных поколений, поможет проанализировать и уточнить популяционно-генетические параметры породы с учетом исторических компонентов ДНК Примечательно, что Бычок (1924 г.р.) отстоит всего на четыре поколения от родоначальника орловского рысака и является рекордистом 1836 года, а Улов (1928 г.р.), также являющийся рекордистом, позволяет оценить изменения, произошедшие через 100 работы с породой. Оба этих жеребца активно использовались в разведении, что позволяет ожидать обнаружения породоспецифичных генетических компонентов у данных представителей породы.
Поэтому цель исследований заключалась в сравнительной характеристике генетических профилей предковых форм орловского рысака и его современных представителей.
2. Материал и методы исследования
Объект исследований. В качестве объекта исследований были выбраны лошади орловской рысистой породы, находящиеся в Алтайском конном заводе и в отделениях Тамбовского ипподрома. Именно поголовье Тамбовского ипподрома обеспечило представительство лошадей разных конных заводов (Воронежская область, Московская область, Пермская область, Республика Чувашия и ряда других), поскольку в связи с реконструкцией Центрального московского ипподрома большая часть поголовья, поступившего из этих заводов, была переведена в Тамбов. Среди исследуемых животных были жеребцы и кобылы, возраст лошадей – 3 года и старше. Все животные прошли оценку по рабочим качествам и экстерьеру. Доля животных, являющихся близкими родственниками, не превышает 5%.
Для получения образцов древней ДНК были использованы скелеты лошадей орловской рысистой породы Бычка (1924 г.р.) и Улова (1928 г.р.), которые хранятся в коллекции Научно-художественного музея Российского государственного аграрного университета – МСХА имени К.А. Тимирязева (рис. 1).

Рисунок 1 - Скелеты и изображения лошадей орловской рысистой породы, используемых для получения древней ДНК
Примечание: а) Бычок (1924 г.р., Молодой Атласный – Домашняя), автор картины И.Н. Раух; б) Улов (1928 г.р., Ловчий – Удачная)
Для получения древней ДНК использовали зубы лошадей: у Бычка был извлечен премоляр, у Улова – нижний клык. Для выделения ДНК из костной ткани применяли набор реагентов COrDIS «ЭКСТРАКТ» ДЕКАЛЬЦИН в соответствии с инструкцией производителя.
Перед проведением лабораторных исследований все образцы были зашифрованы. Анализ проводили по 17 микросателлитных локусов ДНК (AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10, LEX3 и VHL20), рекомендуемых международным обществом генетики животных (ISAG) для лошадей. Определение полиморфизма микросателлитных локусов ДНК лошадей осуществляли в лаборатории биотехнологической компании «ГОРДИЗ» (г. Москва) с использованием 17 микросателлитных локусов, рекомендованных ISAG с использованием собственного набора праймеров (Extra Gene™ DNA Prep 200). ДНК амплифицировали на термоциклере 2720 Cycler Gene Amp PCR (Applied Biosystems Inc., США) с помощью полимеразной цепной реакции (ПЦР) с использованием праймеров с флуоресцентно-меченной меткой. Электрофорез продуктов амплификации осуществлялся на автоматическом 4-капиллярном генетическом анализаторе 3130 DNA Analyzer (Applied Biosystems Inc., США). Расшифровка и документирование полученных графических результатов осуществлялась на компьютере с помощью программного обеспечения автоматической расшифровки результатов фрагментного анализа DNA Genotyper® и Gene Mapper™ ID v3.2.
Статистические методы. Для обработки полученных данных использовали GenAlEx 6.5 для MS Excel
. Для оценки соответствия полученных распределений частот генотипов равновесию Харди-Вайнберга использовали метод хи-квадрат. Значение p<0,05 считалось статистически значимым. Для сравнения генетической структуры исследуемых групп использовался пакет STRUCTURE 2.3.4 как инструмент кластерного анализа . Для математического подтверждения результатов (метод Evanno) результаты, полученные в STRUCTURE 2.3.4, обрабатывались в веб-программе Clampack .3. Результаты исследования и обсуждение
Поскольку лошади орловской рысистой породы могут отличаться в зависимости от региона происхождения, что обусловлено различиями используемых производителей, то мы разделили группу современных лошадей на 6 подгрупп: 1 – лошади из конных заводов Воронежской области (21 гол.), 2 – лошади, в родословной которых встречаются представители разных конных заводов (композитная группа) (36 гол.), 3 – лошади из Московского конного завода (4 гол.), 4 – лошади из конных заводов и хозяйств Украины (9 гол.), 5 – лошади из Пермского конного завода (3 гол.). Отдельную группу 5 составили лошади древней группы. Однако надо отметить, что Бычок, рожденный на конном заводе И. Шишкина, происходит из Воронежской области, а Улов скорее является представителем популяции Московской области.
Оценив результаты генотипирования, было установлено, что в обоих образцах древней ДНК не удалось выявить полиморфизм во всех 17 локусах. Согласно данным А.С. Абдельмановой, если генотипировано менее 50% локусов, то такой образец считается неудачным и должен быть исключен из дальнейшего анализа
. Также были отмечены ошибки амплификации в виде ложных аллелей. В нашем исследовании у Бычка однозначные данные удалось получить для 9 локусов (52.9%), а у Улова – для 12 локусов (70,5%).Оценка результативности генотипирования, проведенная по 17 микросателлитным локусам (табл. 1), показала, что наименьшей успешностью генотипирования отличались локусы HMS7, ASB23, ASB2, HTG10, HTG7, HMS2, CA425.
Таблица 1 - Аллельное богатство лошадей орловской рысистой породы в зависимости от времени и региона происхождения (Na)
Далее мы оценили число приватных аллелей в каждой из групп (табл. 2).
Таблица 2 - Приватные аллели в разных группах лошадей орловской орловской рысистой породыв зависимости от времени и региона происхождения
Интересно, что группе 4 (лошади украинского происхождения) в локусе HMS1, как и у группы 2, также выделен приватный аллель. У лошадей исторической группы приватный аллель Q выделен в локусе VHL20, обладателем которого является самый древний по происхождению Бычок, и который не пересекается с лошадьми других групп.
Для того чтобы оценить, как изменились соотношения генетических компонентов у современных лошадей по сравнению с древними формами, был проведен кластерный анализ с применением ПО STRUCTURE 2.3.4 (рис. 2).
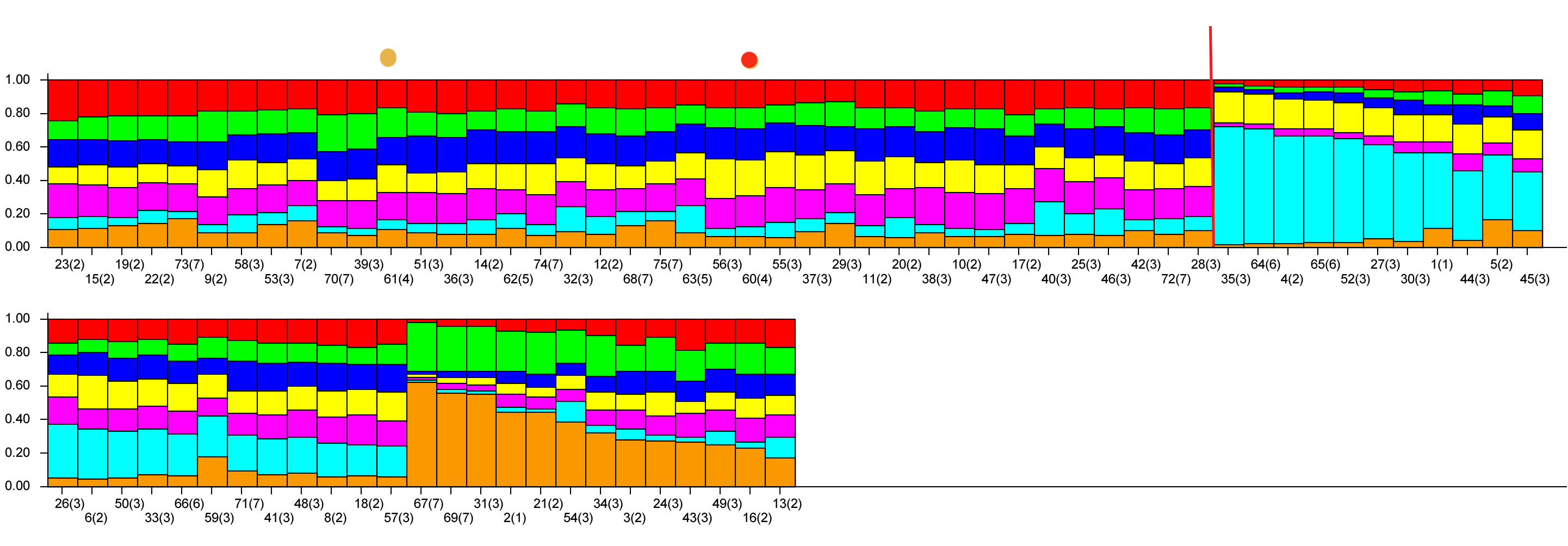
Рисунок 2 - Результаты кластерного анализа лошадей орловской рысистой породы по 17 микросателлитным локусам при К=7 с разбиением по отдельным животным накопленным итогом
Примечание: желтой точкой отмечен профиль Улова, а красной точкой – профиль Бычка
4. Заключение
В настоящей работе представлены результаты генотипирования лошадей орловской рысистой породы по 17 микросателлитным локусам в соответствии с рекомендацией ISAG (International Society for Animal Genetics).
Анализ генетического разнообразия показал, что в породе наблюдается появление внутрипородных групп, которые отличаются по соотношению генетических компонентов. Среди современных представителей породы выявлены животные, генетические профили которых сходны с генетическими профилями древних представителей породы, которые можно считать эталоном породы.
Для понимания механизма появления трех групп лошадей, структура генетических профилей которых сильно отлична от структуры, свойственной орловской рысистой породе, необходимо провести дополнительные исследования, целью которых должен стать поиск причин изменения генетического разнообразия.
Следующим шагом должна быть разработка предложений для Национальной Ассоциации заводчиков и владельцев лошадей орловской рысистой породы по стабилизации генетического разнообразия в породе с помощью корректировки планов племенной работы, а также создании базы данных генетических профилей лошадей орловской рысистой породы.