КОРЕННЫЕ СТАРОВОЗРАСТНЫЕ СЕВЕРОТАЕЖНЫЕ ЕЛЬНИКИ (КРАТКОЕ ГЕОБОТАНИЧЕСКОЕ ОПИСАНИЕ И ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ)
КОРЕННЫЕ СТАРОВОЗРАСТНЫЕ СЕВЕРОТАЕЖНЫЕ ЕЛЬНИКИ (КРАТКОЕ ГЕОБОТАНИЧЕСКОЕ ОПИСАНИЕ И ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ)
Аннотация
Проблема изучения коренных старовозрастных северотаёжных лесов актуальна в связи с исчезновением последних крупных массивов девственной северной тайги. Целью исследования являлось генетико-ботаническое изучение коренных северотаёжных ельников на примере лесного массива, расположенного в северо-западной части Беломорско-Кулойского плато. Выполнено геоботаническое описание коренных северотаежных ельников зеленомошной группы типов леса: черничных и чернично-долгомошных. Установлены основные диагностические признаки эталонных коренных северотаёжных еловых лесов черничного типа. Для коренных северотаёжных еловых лесов черничного типа предложены диагностические виды живого напочвенного покрова. Методом электрофоретического анализа изоферментов исследован генетический полиморфизм ели в северотаёжных популяциях. Установлено, что эти популяции находятся в состоянии, близком к генетическому равновесию с небольшим давлением отбора в пользу гомозигот. Кроме того, они характеризуются стабильно высоким уровнем генетического полиморфизма.
1. Введение
Проблема изучения коренных старовозрастных северотаёжных лесов особенно актуальна в связи с фрагментацией и сведением последних крупных массивов первобытной северной тайги
. При этом само понятие «коренные леса» и его синонимы разными авторами трактуются по-разному . В контексте настоящего исследования под коренными понимаются лесные сообщества, возникшие естественным образом, формированием которых завершается сукцессионный процесс и которые могут, не меняясь, устойчиво функционировать в данных природно-климатических условиях неопределенно долгое время, поскольку полностью адаптированы к этим условиям и в наибольшей степени соответствуют им. Строго говоря, под определение «коренные леса» попадает лишь очень узкий спектр вполне сформировавшихся биоценозов.Очевидно, к коренным следует отнести лесные сообщества, которые сформировались на крайне бедных, сухих, легких по механическому составу почвах и подстилающих породах. В условиях северной тайги это, прежде всего, сосняки лишайниковые и близкие к ним сосняки мохово-лишайниковые и вересково-лишайниковые. Старовозрастные сосняки сфагновой группы, столь широко распространённые в северотаёжной подзоне, по-видимому, можно признать коренными лишь с определёнными оговорками, поскольку эти насаждения не могут быть климаксовыми по определению. В большинстве случаев это промежуточная стадия не пирогенной сукцессии, принимающая состояние зыбкого равновесия в контактной зоне «лес-болото». Высказанное предположение подтверждается тем, что в некоторых крупных водораздельных лесных массивах, перемежающихся огромным количеством разнообразных по флористическому составу верховых и переходных болот, сосняки сфагновой группы могут почти полностью отсутствовать. Вероятно, нельзя признать климаксовыми также широкий спектр травяных и травяно-болотных типов лесных фитоценозов, которые, тем не менее, играют заметную роль в формировании лесных ландшафтов северотаёжной подзоны. Некоторые типы лесных фитоценозов с участием или доминированием лиственницы при выходе к дневной поверхности гипсов и известняков в районах карстовых проявлений, вероятно, правильнее было бы отнести к реликтовым лесам. Сокращение ареала Larix sibirica на русском Северо-Западе началось ещё в атлантическое время в связи с потеплением
. В отличие от восточносибирских насаждений из L. gmelinii и L. cajanderii, сформировавшихся на мерзлотных почвах и полностью адаптированных к местным условиям, насаждения L. sibirica на большей части Европейского Севера являются регрессирующими. В наибольшей же степени определению «коренные леса» в северотаёжной подзоне соответствуют старовозрастные ельники зеленомошной группы, ассоциированные как Empetro-Piceetum obovatae : прежде всего, черничные – Piceetum myrtillosum и близкие к ним чернично-долгомошные, которым присущи все признаки, свойственные состоянию климакса .Целью исследования являлось генетико-ботаническое изучение коренных северотаёжных ельников на примере лесного массива, расположенного в северо-западной части обширного Беломорско-Кулойского плато, в т.н. бассейне малых рек (водораздел рек Мудьюга и Кадь). В указанном районе никогда не осуществлялась никакая хозяйственная деятельность, а его удалённость от дорог исключала до последнего времени сколько-нибудь заметное антропогенное воздействие. Учитывая чрезвычайно развитую гидрографическую сеть и ничтожную вероятность сухих гроз, не исключено, что этот массив формировался, не претерпевая каких бы то ни было катастрофических изменений в течение длительного времени. Сказанное позволяет отнести старовозрастные ельники указанного района к эталонам тёмнохвойных северотаёжных лесов.
2. Методы и принципы исследования
Геоботаническое описание фитоценозов проводили в соответствии с общепринятыми методами исследований
. При обработке материалов полевых исследований степень проективного покрытия для каждого вида в границах пробной площади определялась по результатам 30-ти повторностей и переводилась затем в баллы по 6-бальной шкале. Класс встречаемости для каждого вида определялся из отношения числа учётных площадок, на которых было отмечено присутствие данного вида к их общему числу, выраженному в процентах. Далее этот показатель также переводился в баллы по 5-бальной шкале. Константность вида определялась как произведение балла проективного покрытия на коэффициент (класс) встречаемости. Обилие эпифитных лишайников оценивалось визуально по 5-бальной шкале, при этом подразумевалась степень обилия эпифитов на нижних боковых ветвях и стволах деревьев до высоты 1/3 h. Наличие сухостоя и степень захламлённости участка (свежего валежа) также оценивались визуально по 5-бальной шкале.Изучение генетического полиморфизма природных популяций ели проводили методом электрофоретического анализа изоферментов
. На основе результатов многолетних исследований были выбраны те ген-ферментные системы, которые проявляли полиморфизм при электрофоретической разгонке и дававшие стабильные спектры при инкубировании гелевых пластин. Для электрофореза использовали гомогенаты эндоспермов. С целью сокращения объёмов лабораторных исследований совместно гомогенизировали 10-12 эндоспермов отдельного дерева. При этом на электрофореграмме фактически проявлялся генотип данного дерева и исключалось появление межлокусных гетеродимеров, характерных для соматических тканей, что облегчало последующую дешифровку.3. Результаты исследования и обсуждение
Геоботанический профиль (рис. 1) был заложен в северо-западной части Беломорско-Кулойского плато, на водоразделе рек Кадь и Мудьюга, в их верхнем и среднем течениях.
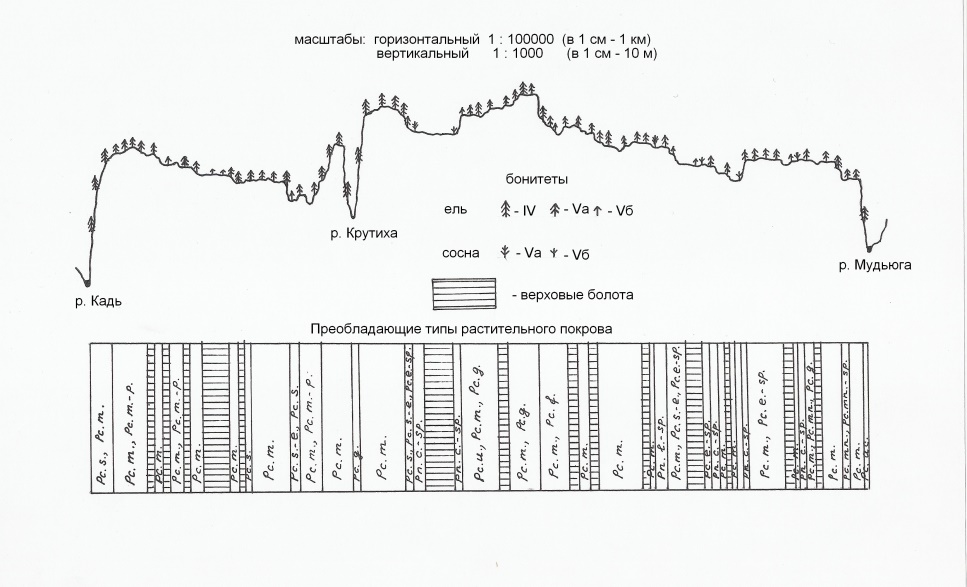
Рисунок 1 - Мозаика северотаежных ассоциаций на водоразделе рек Мудьюга и Кадь
Примечание: Ельники: Pc.m. – Piceetum myrtillosum (ельник черничный); Pc.m.-p. – Pyceetum myrtilloso-polytrichosum (ельник чернично-долгомошный); Pc.g. – Piceetum gymnocarposum (ельник разнотравный); Pc.s. – Piceetum spinulosum (ельник папоротниковый); Pc.s.-e. – Piceetum spinuloso-equisetosum (ельник хвощево-папоротниковый); Pc.e. – Piceetum equietosum (ельник хвощевый); Pc.e.-sp. – Piceetum equisetoso-sphagnosum (ельник хвощево-сфагновый); Pc.mn.-sp. – Piceetum menyanthoso-sphagnosum (ельник вахто-сфагновый); Pc.mn. – Piceetum menyanthosum (ельник вахтовый); Pc.f. – Piceetum filipendulosum (ельник таволговый); Pc.u. – Piceetum uliginosum (ельник голубично-кустарничковый); Pc.ac. – Piceetum aconitosum (ельник широкотравный). Сосняки: Pn.c.-sp. – Pinetum caricoso-sphagnosum (сосняк осоково-сфагновый); Pn.l.-sp. – Pinetum ledoso-sphagnosum (сосняк багульниково-сфагновый)
Примеры описаний старовозрастных северотажных ельников зеленомошных в несколько сокращённом виде даны в табл. 1.
Таблица 1 - Геоботаническая характеристика коренных северотаёжных еловых лесов зеленомошной группы (ельники черничные свежие и влажные)
№ описания | 8 | 9 | 11 | 12 | 13 | Константность | Класс встречаемости | |||||
Количество сосудистых видов | 19 | 17 | 17 | 18 | 16 | |||||||
Древостой | ||||||||||||
Состав древостоя | 10Е+Б | 10Е+Б | 10Еед.Б | 9Е1Б | 7Е3Б | |||||||
Сомкнутость крон | 0,7 | 0,7 | 0,6 | 0,7 | 0,6 | |||||||
Степень захламлённости | 4 | 3 | 4 | 5 | 2 | |||||||
Наличие сухостоя | 4 | 4 | 4 | 3 | 3 | |||||||
Обилие эпифитов | 3 | 3 | 4 | 4 | 3 | |||||||
Подрост и подлесок | ||||||||||||
ОПП, % | 4 | 4 | 5 | 5 | 5 | |||||||
| Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости |
|
|
Picea abies x P. obovata | 2 | - | 2 | - | 2 | - | 2 | - | 2 | - | - | V |
Sorbus aucuparia | 1 | - | 2 | - | 2 | - | 2 | - | 2 | - | - | V |
Betula pubescens | 1 | - | 1 | - | 1 | - | 1 | - | 1 | - | - | V |
Juniperus communis | 1 | - | - | - | 1 | - | - | - | II | |||
Populus tremula | - | - | - | - | 1 - |
| - | I | ||||
Травяно-кустарничковый ярус | ||||||||||||
ОПП, % | 58 | 84 | 69 | 61 | 67 | |||||||
| Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости |
|
|
Vaccinium myrtillus | 4 | 5 | 5 | 5 | 4 | 5 | 4 | 5 | 4 | 5 | 21,0 | V |
Lerchenfeldia flexuosa | 2 | 5 | 1 | 1 | 2 | 5 | 3 | 4 | 3 | 5 | 9,6 | V |
Vaccinium vitis-idaea | 2 | 2 | 2 | 3 | 2 | 5 | 2 | 3 | 3 | 5 | 8,2 | V |
Linnea boreales | 2 | 3 | 1 | 2 | 1 | 1 | 2 | 5 | 2 | 3 | 5,0 | V |
Chamaepericlymenum suecicum | 2 | 2 | 2 | 3 | 3 | 4 | 1 | 1 | - | 4,6 | IV | |
Trientalis europae | 2 | 3 | - | 1 | 2 | 2 | 3 | 1 | 1 | 3,0 | IV | |
Luzula pilosa | 2 | 2 | 1 | 1 | - | 2 | 2 | 2 | 3 | 3,0 | IV | |
Empetrum nigrum | 1 | 1 | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 2,2 | V |
Vaccinium uliginosum | 1 | 1 | 2 | 1 | 2 | 2 | - | 2 | 1 | 1,8 | IV | |
Equisetum sylvaticum | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 1 | - | 1,6 | IV | |
Lycopodium annotinum | 1 | 1 | - | 1 | 1 | 1 | 1 | 2 | 2 | 1,4 | IV | |
Gymnocarpium dryopteris | 2 | 1 | - | - | 2 | 2 | - | 1,2 | II | |||
Rubus chamaemorus | - | 2 | 3 | - | - | - | 1,2 | I | ||||
Melampyrum sylvaticum | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1,0 | V |
Majanthemum bifolium | - | - | - | 1 | 3 | 1 | 2 | 1,0 | II | |||
Ramischia secunda | 1 | 1 | 1 | 1 | 1 | 1 | - | - | 0,6 | III | ||
Carex praecox | - | 1 | 1 | 1 | 1 | - | - | 0,4 | II | |||
Solidago virgo-aurea | - | - | - | - | 2 | 1 | 0,4 | I | ||||
Dryopteris spinulosa | - | - | - | 2 | 1 | - | 0,4 | I | ||||
Monesis uniflora | - | 1 | 1 | 1 | 1 | - | - | 0,4 | II | |||
Deschampsia caespitosa | 1 | 1 | - | - | - | - | 0,2 | I | ||||
Мохово-лишайниковый ярус | ||||||||||||
ОПП, % | 78 | 88 | 74 | 76 | 66 | |||||||
| Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости | Балл проективного покрытия | Класс встречаемости |
|
|
Pleurozium Schreberii | 4 | 4 | 3 | 4 | 3 | 5 | 3 | 5 | 3 | 5 | 14,6 | V |
Hylocomium splendens | 3 | 4 | 3 | 3 | 3 | 4 | 3 | 4 | 3 | 4 | 11,4 | V |
Dicranum scoparium | 3 | 3 | 3 | 5 | 3 | 3 | 3 | 3 | 3 | 3 | 10,2 | V |
Polytrichum commune | 3 | 5 | 3 | 2 | 2 | 3 | 3 | 4 | 1 | 1 | 8,0 | V |
Barbilophozia lycopodioides | 1 | 1 | 2 | 1 | 3 | 4 | 2 | 2 | 3 | 5 | 6,8 | V |
Dicranum polysetum | 2 | 1 | - | 2 | 2 | 2 | 2 | 3 | 3 | 3,8 | IV | |
Sphagnum fimbriatum | - | 4 | 2 | 3 | 3 | 2 | 1 | - | 3,8 | III | ||
Ptilium crista-castrensis | 3 | 2 | - | 1 | 1 | 3 | 3 | 2 | 1 | 3,6 | IV | |
Sphagnum compactum | 3 | 1 | - | - | 2 | 1 | - | 1,0 | II | |||
Cladonia sylvatica | - | - | 1 | 1 | 1 | 1 | 2 | 1 | 0,8 | III | ||
Rhytidiadelphus triquetrus | 1 | 1 | - | - | 1 | 1 | - | 0,4 | II | |||
Sphagnum warnstorfii | - | - | - | 2 | 1 | - | 0,4 | I | ||||
Cladonia difformis | 1 | 1 | - | - | 1 | 1 | - | 0,4 | II | |||
Cladonia rangiferina | - | - | 1 | 1 | - | 1 | 1 | 0,4 | II | |||
Cladonia elongata | - | - | - | 1 | 1 | - | 0,2 | I | ||||
Cladonia fimbriata | - | - | - | 1 | 1 | - | 0,2 | I | ||||
Peltigera aphthosa | - | - | - | 1 | 1 | - | 0,2 | I | ||||
Участки №№ 8, 11 и 12 – типичные ельники черничные свежие. Участок № 9 – переходный от ельника черничного к ельнику чернично-долгомошному. Участок № 13 – ельник черничный свежий, не исключена вероятность того, что не позднее 1740-1750 г. этот участок был пройден низовым пожаром.
Все древостои абсолютно разновозрастные с явным преобладанием Picea abies x P. obovata с небольшой примесью Betula pubescens: состав древостоев от 10Еед.Б до 7Е3Б, средний состав: 10Е+Б. Возраст деревьев ели в первом ярусе варьирует от 100 до 280 лет и составляет, в среднем, 180-220 лет. Возраст березы составляет, в среднем, 80-100 лет. Относительно низкопродуктивные: IV (V) классов бонитета насаждения, среднеполнотные (0,6-0,7).
В подросте доминирует ель с небольшой примесью березы, очень редко – осины. Состав подроста варьирует от 7Е3Бед.Ос до 9Е1Б и составляет, в среднем 8Е2Б. Количество подроста ели от 1800 до 5200 шт./га, березы – в среднем 700 шт./га, осины – не более 50 шт./га. Крупный и средний подрост ели распределен по площади более или менее равномерно, мелкий – часто приурочен к микроповышениям, особенно излюбленные места для него – полуразложившийся еловый валеж. Подрост березы распределён равномерно-рассеянно, но часто тяготеет к «окнам» в пологе древостоя. В подлеске явно доминирует рябина, изредка встречается можжевельник. Общая сомкнутость (густота) подлеска, как правило, невысока и не превышает 5-10%.
В травяно-кустарничковом ярусе явно доминирует Vaccinium myrtillus, проективное покрытие её местами достигает 100% и варьирует по участкам, в среднем, от 40 до 60%. Высота Vaccinium myrtillus достигает местами 60-70 см и составляет в большинстве случаев 30-40 см, высота определена приблизительно, поскольку часто кустарничек принимает стелющиеся формы. Vaccinium myrtillus повсеместно сопутствует V.vitis-idaea, распределение последней по площади рассеянно-равномерное, на хорошо освещённых и прогреваемых кочках этот кустарничек может доминировать. Высота его значительно меньше первого и в целом он явно соподчинен ему. Повсеместно, по большей части тяготея к микровпадинкам на куртинках-подушках сфагнумов и кукушкина льна произрастает V. uliginosum. Empetrum nigrum, являясь ксеро-мезо-гигрофитом также отмечена повсеместно в небольшом количестве: 0,5-1,5%, и лишь в более влажных местообитаниях (черничный влажный тип) её общее проективное покрытие может достигать 5%. Задернение в ельниках черничных практически отсутствует. Lerchenfeldia flexuosa отмечена повсеместно, но её размещение равномерно-рассеянное, а общее проективное покрытие не превышает 6%, она явно соподчинена, о чем свидетельствует отсутствие генеративных органов, и крайне редко образует дернинки. Deschampsia caespitosa отмечена единично и не является типичным видом для данного типа леса. Повсеместно, иногда достаточно обильно, особенно по микроповышениям встречается Linnea boreales, и, учитывая размеры этой лианы, необходимо обратить внимание на её изобилие в коренных северотаёжных ельниках черничных. Повсеместно, иногда достаточно обильно, встречаются типичные лесные виды таёжного мелкотравья: Chamaepericlymenum suecicum, Trientalis europae, Gymnocarpium dryopteris, Majanthemum bifolium, Ramischia secunda. Повсеместно, но все же чаще приуроченно к более увлажнённым местообитаниям, встречается Equisetum sylvaticum. Повсеместно рассеянно в небольшом количестве (проективное покрытие 0,2-1,2%) встречается Luzula pilosa. Monesis uniflora также, по-видимому, распространена повсеместно, но, учитывая размеры растения, отмечена не везде. Dryopteris spinulosa отмечен единично, поскольку тяготеет к более увлажненным и богатым почвам и не характерен для ельников черничных. Практически повсеместно, часто по микроповышениям отмечено присутствие Lycopodium annotium, который, разрастаясь, может занять компактно небольшое пространство, не проявляя, однако, экспансии. Melampyrum sylvaticum распространён повсеместно-рассеянно, но в среднем его общее проективное покрытие редко превышает 0,1-0,2%, и он редко образует генеративные органы вследствие недостатка освещенности. Solidago virgo-aurea приурочен к наиболее освещённым участкам – «окнам» и встречается более редко. Само растение обычно представляет розетку листьев, не формируя цветоносного стебля, что также свидетельствует о недостатке освещённости. В наиболее влажных местах обитания и при переходе к более увлажнённым типам леса встречается Rubus chamaemorus.
Характерной особенностью ельников черничного типа является хорошо сформировавшийся мохово-лишайниковый ярус
. В напочвенном покрове преобладают зелёные листостебельные мхи: Pleurozium Schreberii, Hylocomium splendens, Dicranum scoparium, Ptilium crista-castrensis, Dicranum polysetum. Повсеместно, но неравномерно, встречаются Polytrichum commune (общее проективное покрытие варьирует, в среднем, от 0,4 до 11,1%), он может произрастать как в виде подушек-дернинок, так и рассеянно-одиночно, а также некоторые виды сфагновых мхов: Sphagnum fimbriatum образует ярко-зелёные рыхлые высокие дернинки, а Sph. compactum – мелкие, плотные, желто-зелёные подушки. Непосредственно в напочвенном покрове явно доминируют Pleurozium Schreberii (ОПП 15-25%) и Hylocomium splendens (ОПП 10-17%), которые в более затенённых и увлажнённых местообитаниях замещаются печеночником Barbilophozia lycopodioides, степень проективного покрытия которого может достигать 15%. Ptilium crista-castrensis более приурочен к микроповышениям – полуразложившемуся валежу и полуистлевшим пням и встречается не так обильно (ОПП 0,5-5%). Dicranum scoparium (ОПП 10-20%) и Dicranum polysetum (ОПП 1-5%) совместно с успехом произрастают как на почве, так и по микроповышениям. Редко встречается Rhythydiadelphus triquetrus, который более типичен для сырых, относительно богатых почв ручейных, травяных и травяно-болотных типов леса. Изредка на почве встречаются вкрапления кустистых кладоний: Cladonia sylvatica и произрастающей совместно с ней, но в меньших количествах Cl. rangiferina. На высоких пнях и старом валеже повсеместно встречаются многие виды бокальчатых кладоний: Cladonia difformis, Cl. elongata, Cl.fimbriata и др., а также различные виды р. Peltigera.В целом коренные северотаёжные ельники черничного типа характеризуются хорошо развитым травяно-кустарничковым (ОПП 58-84%) и мохово-лишайниковым (ОПП 66-88%) ярусами и относительно небогатой (15-20) флорой сосудистых видов. Достаточно обильны эпифитные лишайники, среди которых повсеместно доминирует Hypogymnia physodes, обычно произрастающая на нижних ветвях ели и стволах березы, а также бородатые лишайники: Usnea barbata, U.dasypoga, Bryopogon sp., Evernia sp., и др. Несколько реже встречаются различные виды р. Parmelia: P. сaperata, P. сentrifugata и др. Основания стволов ели часто густо заселены лепрозными формами Lichenes imperfectii. Для всех старовозрастных северотаёжных ельников характерна повышенная фаутность деревьев и высокая степень риска ветровала, особенно в очагах корневой губки.
Анализ генетического полиморфизма ели в коренных северотаёжных популяциях дан в табл. 2.
Таблица 2 - Показатели генетического полиморфизма северотаёжных популяций ели
Показатели | Популяция | ||||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | |
Наблюдаемая гетерозиготность, Но, % | 16,4 | 26,4 | 30,6 | 18,8 | 18,8 | 20,2 | 11,8 |
Ожидаемая гетерозиготность, Не, % | 25,4 | 29,6 | 28,6 | 28,0 | 28,4 | 28,3 | 22,7 |
Среднее число аллелей на локус, µ | 2,4 | 2,4 | 2,1 | 2,3 | 2,4 | 2,2 | 2,3 |
Коэффициент инбридинга, F | 0,290 | 0,185 | 0,170 | 0,255 | 0,296 | 0,286 | 0,347 |
Популяции под №№ 1-6 географически идентичны описанным выше участкам и соответствуют коренным ельникам черничного и долгомошного типов. Популяция под № 7 находится в Усинском лесхозе республики Коми. Структура генофондов коренных ельников черничного типа близка на всей изученной территории. Эти популяции находятся в состоянии, близком к генетическому равновесию с небольшим давлением отбора в пользу гомозигот. Кроме того, они характеризуются стабильно высоким уровнем генетического полиморфизма.
4. Заключение
Основными диагностическими признаками эталонных коренных северотаёжных еловых лесов черничного типа являются:
1. Абсолютное доминирование Picea abies (L.) Karst. x P. obovata (Ledeb.);
2. Абсолютная разновозрастность Picea abies (L.) Karst. x P. obovata (Ledeb.), отсутствие чётко выраженных возрастных поколений;
3. Хорошо развитый, почти сплошной живой напочвенный покров, в котором доминируют ягодные кустарнички (Vaccinium myrtillus) и зелёные листостебельные мхи;
4. Относительно бедный (15-20 видов) состав сосудистой флоры;
5. Часто выраженная мозаичность напочвенного покрова, обусловленная неоднородностью почвенно-гидрологических условий;
6. Стабильный видовой состав дереворазрушающих грибов: Fomitopsis annosa (Fr.) Karst., Armillariella mellea (Vahl. ex Fr.) Karst., Phellinus pini var. abietes (Karst.) Pil., Polystictus circinatus var. triqueter Bres., Fomitopsis pinicola (Sw. et Fr.) Karst., Inonotus obliquus (Pers.) Pil., Fomes fomentarius (L.) Gill., Piptoporus betulinus (Bull. ex Fr.) Karst., Phellinus igniarius (L. ex Fr.) Quel и некоторые др.;
7. Богатая энтомофауна ксилофагов;
8. Чрезвычайно частая встречаемость жилищ Formica rufa, высокая плотность поселений древесных видов формицидов;
9. Высокий уровень генетического полиморфизма, генетическая стабильность и самодостаточность сформировавшихся популяций Picea;
Вся совокупность перечисленных признаков старовозрастных северотаежных ельников обусловлена очень длительным периодом формирования этих популяций. Минимальная продолжительность формирования разновозрастной популяции ели, по мнению Г.П. Морозова , составляет 550 лет. По-видимому, в северотаёжной подзоне этот срок может превышать 1100-1200 лет. Роль пирогенного фактора, как причины катастрофических метаморфозов в отношении еловых лесов, по-видимому, сильно преувеличена и не исключено, что в ряде случаев эталонные ельники формировались эволюционно с начала отступления ледника, т.е. в течение последних 9-11 тыс. лет.
При описании фитоценозов важным шагом является установление диагностических видов. В отношении северотаёжных лесных сообществ используемые большинством геоботаников формальные методы анализа табличных материалов (метод Браун-Бланке и др.) малоприемлемы, т.к. эти сообщества характеризуются крайне скудным видовым составом. Многие типы леса различаются лишь соотношением одних и тех же видов, часто не столь существенным. В нашем понимании, под диагностическими понимаются виды, характерные для данных местообитаний и присущие чаще всего и в наибольшей степени только им. Для коренных северотаёжных еловых лесов черничного типа таким диагностическим видом является Barbilophozia lycopodioides, а также другие виды печеночников из рр. Lophozia и Barbilophozia.